紅檜與台灣扁柏
![]()
![]()
![]()
![]()
![]()
紅檜與台灣扁柏為台灣地區珍貴之針葉樹,在分類學上隸屬於松柏目(Coniferae),柏科(Cupressaceae),扁柏屬。此屬樹種全球的分佈僅有7種,除台灣地區兩種外,其餘包括日本產2種為日本扁柏(Chamaecyparis obtusa)及日本花柏(C.pisifera);美國產3種為羅生柏(C. lawsoniana)、拿加遜扁柏(C. nootkatensis)及側葉扁柏(C. thyoides)。這7種樹木均為巨大之喬木,且有相當的經濟價值。
紅檜與台灣扁柏木材鑑別特徵
外觀特徵 |
||
| 樹名 | 紅檜 | 台灣扁柏 |
| 邊心材 | 邊心材區別稍明顯,心材率較低 | 邊心材區別稍明撇i,心材率較高 |
| 材色 | 邊材為淡黃灰白色·心中才為紅黃到淡紅棕色 | 邊材為黃白色,心材為淡紅棕色到黃棕色 |
| 年輪 | 年輪明顯,年輸寬較寬,早材到晚材的變化是漸進的 | 生長年輪明顯,寬度極狹小,橫斷面常有小龜裂情形,早材到晚材的變化是突然的 |
| 木理 | 通直,弦切面美麗花紋,不具密而細鋸齒狀木紋 | 通直,弦切面具密而細鋸齒狀木紋 |
| 觸感 | 木肌細緻 | 木肌細緻,富光澤 |
| 氣味 | 香氣強,不具辛辣味 | 芳香,具辛辣味 |
| 正常樹脂溝 | 不具有 | 不具有 |
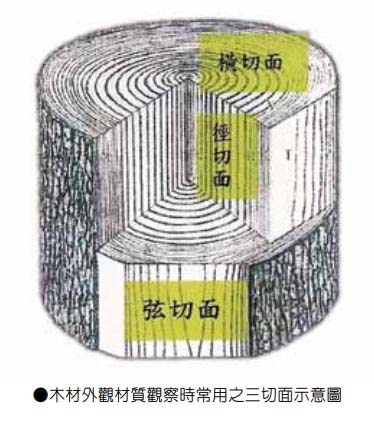

台灣紅檜橫切面

台灣扁柏橫切面
台灣紅檜


台灣紅檜徑切面與弦切面
英文名:Taiwan red cypress
學 名:Chamaecyparis formosensis
科 名:柏木科 Cupressaceae
產 地:台灣
說明:木材外觀與台灣扁柏略相似,唯紅檜色澤較帶淡紅色。其加工性質大致與台灣扁柏相似,生材比重較台灣扁柏大,氣乾材卻較台灣扁柏小,耐蟻性與耐濕性則較台灣扁柏強。 為老樹常具有蓮藕朽,俗稱為蓮根材,致使降低木材之利用價值,但可用於雕刻用材。於台灣市場上與台灣扁柏通稱為檜木。
木材特徵:
1. 邊心材區別明顯。
2. 年輪明顯,春材向秋材漸進移行而分明。
3. 木材具柏木香氣。
用途:建築、家具、棺木、雕刻等。
台灣扁柏


台灣扁柏徑切面與弦切面
英文名:Taiwan yellow cypress
學 名:Chamaecyparis taiwanensis
科 名:柏木科 Cupressaceae
產 地:台灣
說明: 台灣扁柏為台灣產木材中最優良之木材, 木理通直均勻,木肌細緻,富光澤,木材具芳香及辛辣味。木材輕軟中庸,富彈性,耐腐性及耐蟻性,乾燥容易且乾燥狀態極良好,少反翹變形,收縮極小易割裂,刨削加工容易,刨面極光滑而精緻,釘著性良好,吸漆性佳,塗裝後更顯精良。
木材特徵:
1. 邊心材區別明顯。但界限不明顯,邊材淡紅黃白色,心材淡紅黃色至黃褐色
2. 年輪極狹,春秋材漸進移行
3. 木材具柏木香氣。
用途:建築、家具、棺木、雕刻等。
紅檜與台灣扁柏材質
木材為一種由不同種類之橫向與縱向細胞組合而成的有機性生物材料,由於遺傳特性、環境因子及經營措施的不同,會使得木材組成細胞的種類、比例及化學組成分產生差異,致使木材各項基礎性質存在變異的情形。由於木材之組成複雜,各項材質不僅會因樹種不同而有異,即便同一樹體內不同部位的材質,亦可能會有極大的差別,故要有效利用檜木資源,必須對檜木的基礎材質有充分的瞭解。
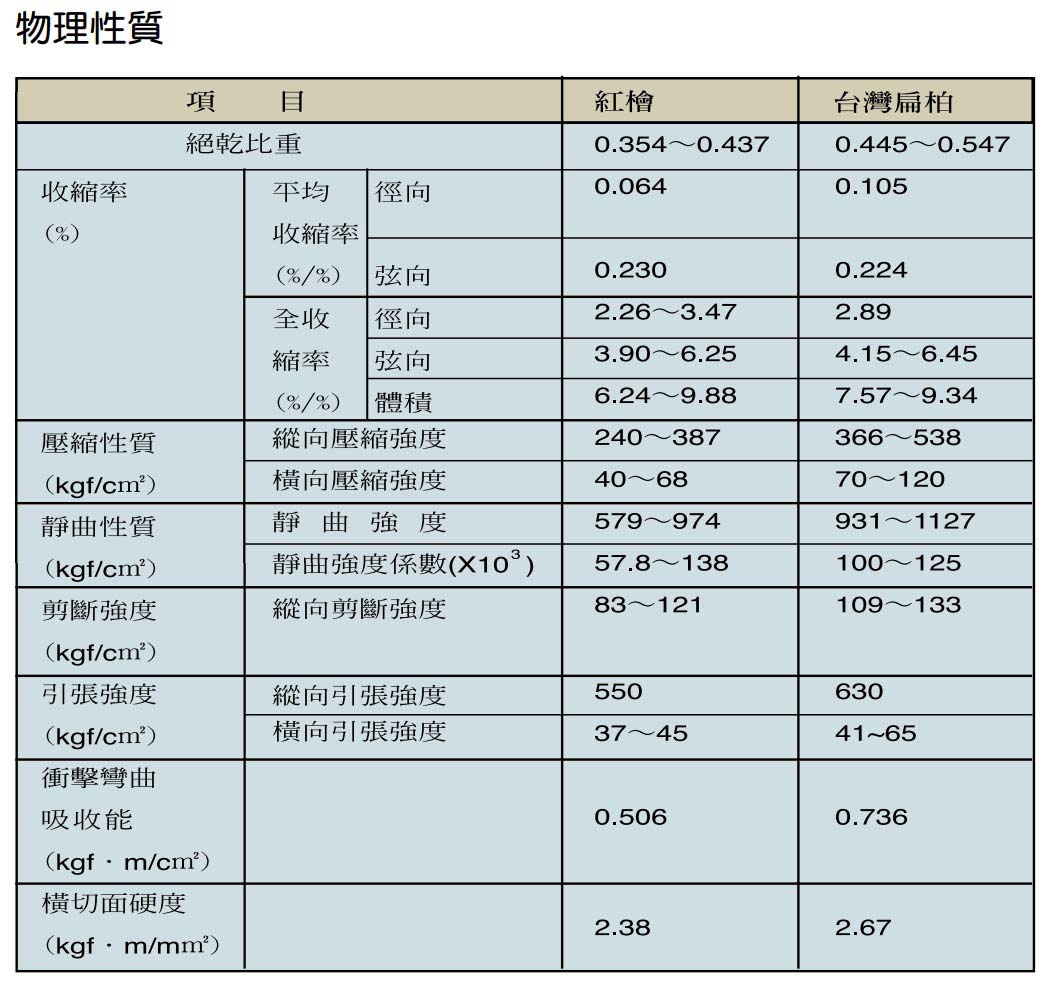
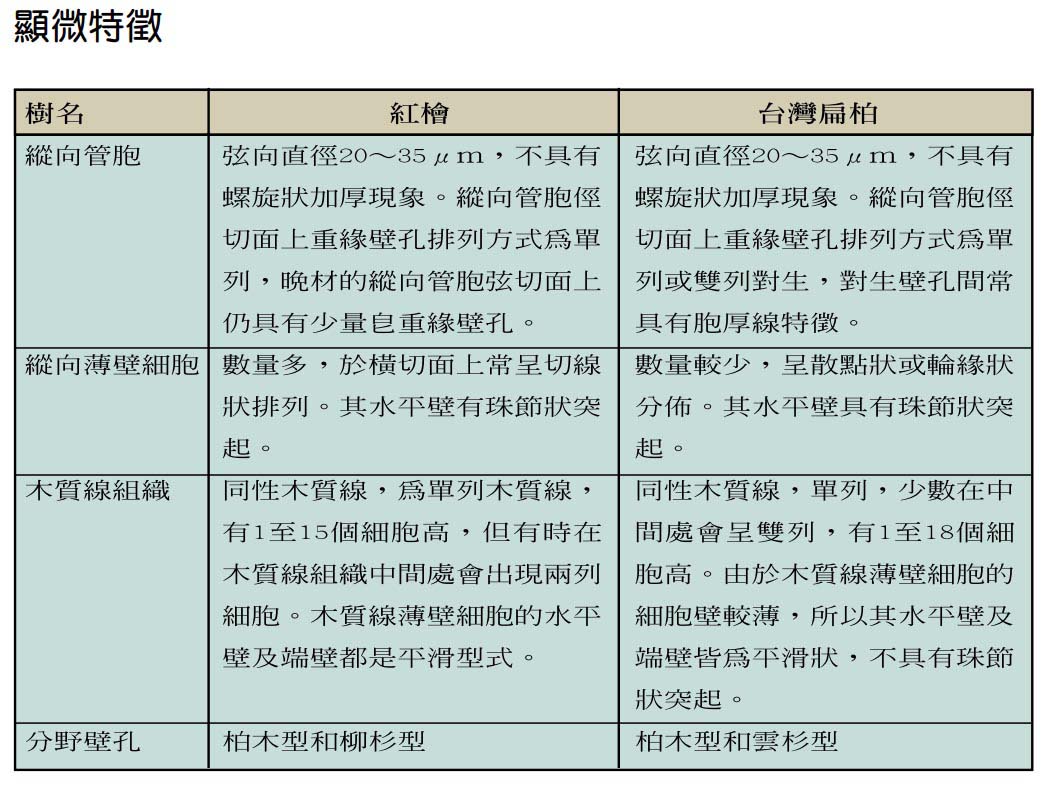
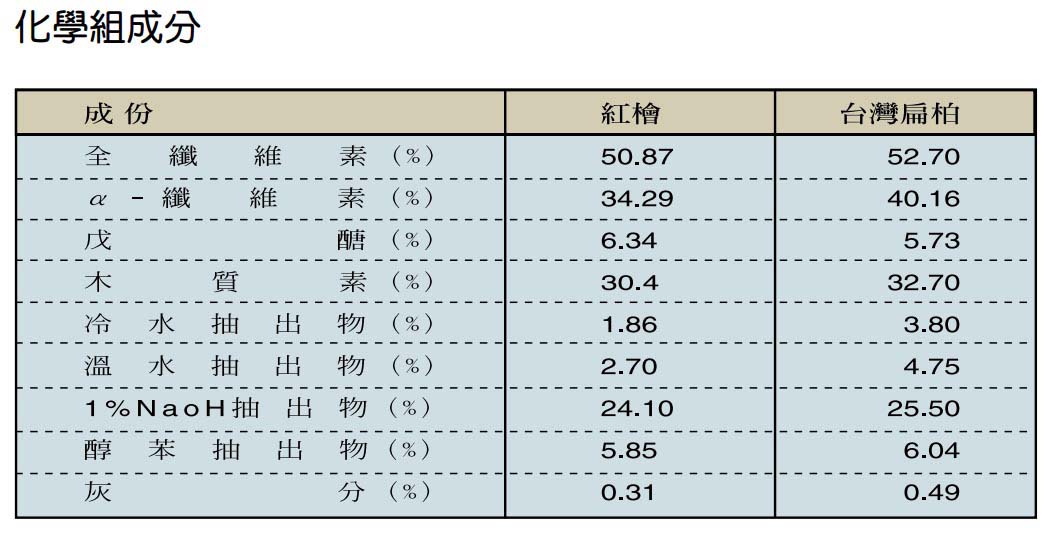
檜木的生態習性
檜木分布
檜木屬於扁柏屬植物,性喜溫溼,主要分布在北半球中緯度之太平洋西側及北美洲東西兩岸等處。由抱粉學的研究推測,檜木林600萬年前普遍存在地球各處,隨著物種的演化,檜木逐漸被被子值物取代。更由於冰河期的結束,地球環境變化的結果,使得全球僅剩下7種檜木,群落分佈於台灣、日本、美國西北海岸及東南海岸。
從扁柏屬植物親緣演化研究的結果顯示,扁怕族群中的側葉扁柏最旱被分歧而出,羅生扁怕、日本扁柏及台灣扁怕形成一分枝群,紅檜、日本花柏則形成另一分枝群,而拿加遜扁怕與其他扁柏族群之親緣關係則較認疏遠,生態演化的研究顯示,扁怕族群可能發源於北美,經由日本群島再遷徙至台灣,分類上有將台灣扁怕歸類認日本扁柏一變種的看法。
由台灣扁怕及紅檜族群遺傳變異的研究,台灣扁怕與紅檜的葉綠體DNA不具分化狀況,這些推論指出台灣檜木族群可能是原有大族群,歷經冰河期後所殘留之小族群,未必是自冰河退卻後花粉錢散所提供之基因交流頻繁所造成。以逢機擴增多型性DNA分子標誌研究族群分化,顯示出棲蘭山之台灣扁怕與紅檜族群均較阿里山族群有較高之遺傳變異,可能與棲蘭山為大量紅檜及台灣扁柏之群生地有關。台灣檜木分布於台灣上要山脈兩側中高海拔山區之雲霧林帶,天然林社會組成結構,上層以檜木認優勢,多與其他針葉樹種混生,如台灣杉、巒大杉及鐵杉等。次層以樟科及殼斗科為主之闊葉樹種,再下之灌木層由灰木科、杜鵑花科、茶科值物等組成之針闊葉樹林,可分認台灣扁柏亞林型及紅檜亞林型。台灣扁怕分布北限大致在北插天山一帶,南限則在阿里山、玉山秀姑巒一帶;紅檜北限亦為北插天山一帶,(但因分布海拔較低,所以分布地較台灣扁柏略認偏北),南限則至大武山一帶,(則又比台灣扁怕分布更南)由於二者生態習性略有差異,分布上素有北(扁柏)南檜(紅檜)之謂。
由於日本人於1912年起在台灣大規模伐採檜木,加上日式建築風格及工法之引進,帶動台灣對檜木之使用與重視。
台灣於民國79年10月禁伐一級木(檜木林),80年11月全面禁止砍伐天然林後,檜木益顯珍貴。
由於檜木材源的日漸稀少,天然檜木林巨木也因環境生態保育之故而不再開發;有感於天然檜木為台灣的珍貴資源,而檜木的開發亦曾為台灣經濟嶺展上重要的環節之一。
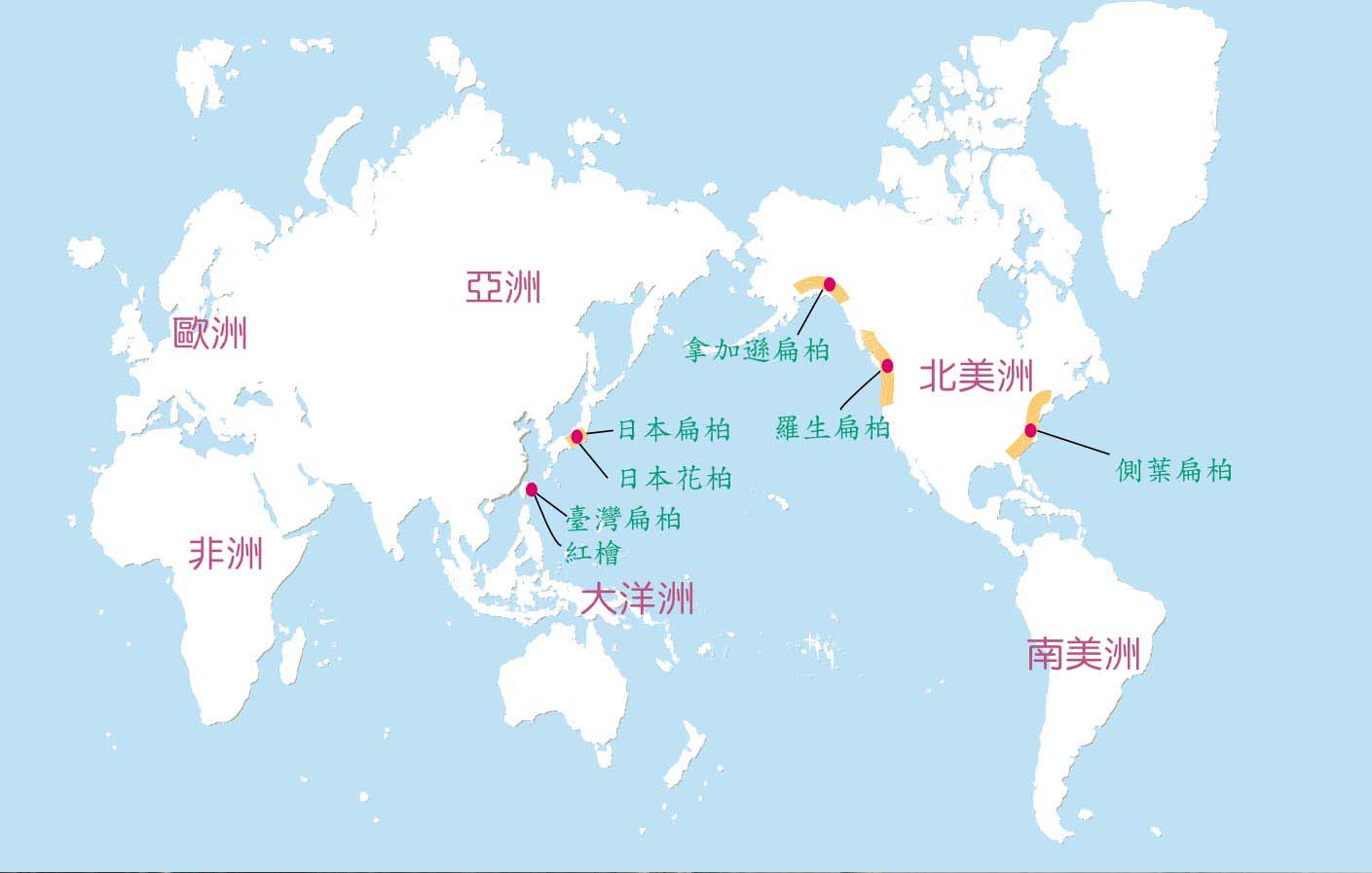
世界檜木之種類與分佈地區
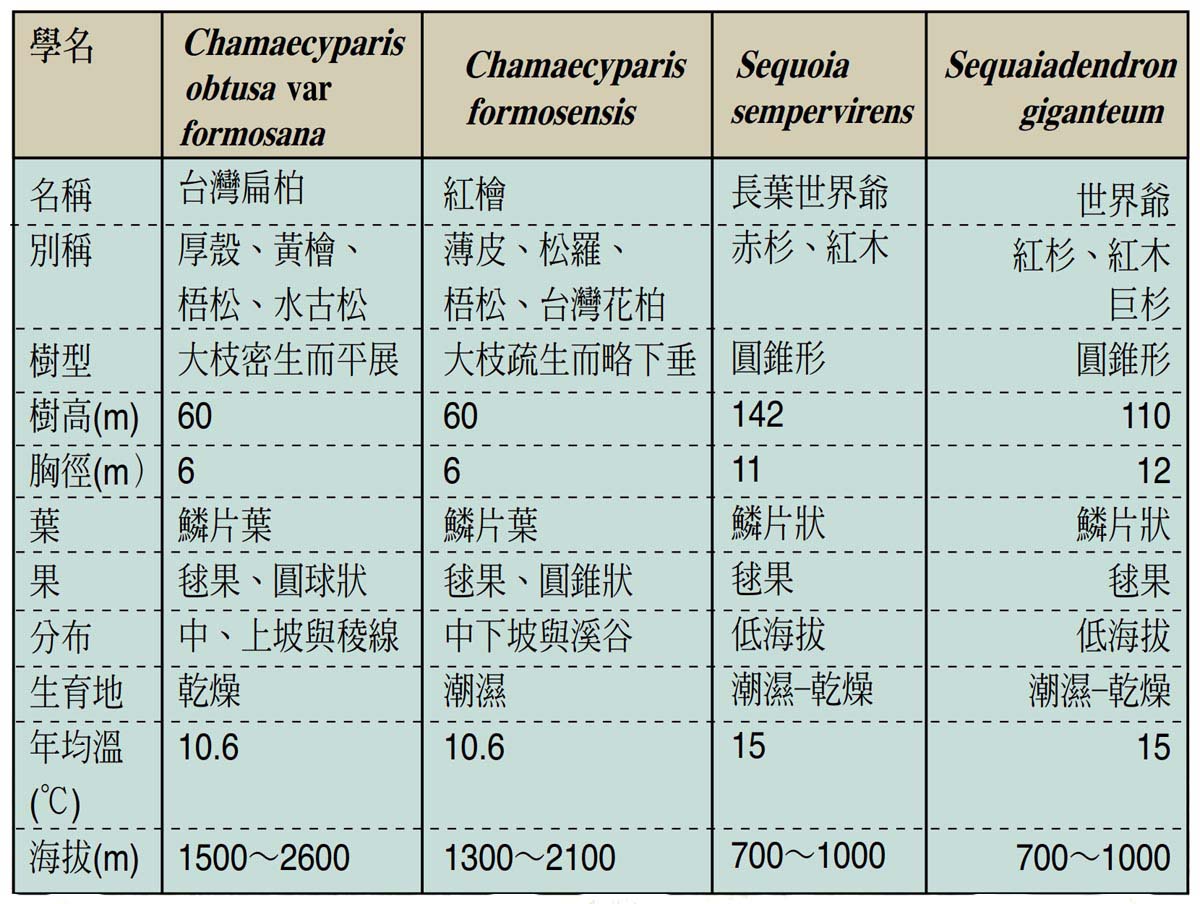
全球最出名之巨木當推長葉世界爺及世界爺為最,此二樹種多生長於美國西部太平洋沿岸中低海拔之潮濕山區,均有著巨大之胸徑與樹高。台灣之巨木以紅檜及台灣扁柏最著名,針對紅檜與台灣扁柏之樹形、各部尺寸、生長環境等與世界爺及長葉世界爺比較之結果,以表列於下:
地球上活的最久的樹
在非洲西部加那利亞島上有棵龍血樹,樹齡高達8000多歲,可惜的是在1868年毀于一場風災,現存於地球上被認定為最老的活樹,則是在美國加州被發現的刺毬果松。此株刺毬果松在1954年被發現生長在加州白山海拔3000公尺全無水源分佈的礫石山區,在艱惡的生長環境下,樹體僅有少數綠葉,其他部分則視之如死木,刺毬果松以如此的生存策略存活於條件極差之環境下。

非洲西部加那利亞島上龍血樹。

地球上活的最久的樹─美國加州被發現的刺毬果松。
地球上最高、最大的樹
世界上最大的樹世界爺一巨杉,和世界上最高的樹長葉世界爺一赤杉,都生長在美國西部的加利福尼亞州,國家公園的巨杉及赤杉林中,超過110公尺樹高的大樹比比皆是。
澳洲的杏仁按樹林,皆為樹高百公尺以上之巨木,其中最高者高達156公尺,相當於50層的高樓,為現今所發現地球上最高之樹體,次高者則為生長在美國高142公尺的長葉世界爺。台灣最高的樹種則為台灣杉,通常散生於檜木林中,根據林務局進行台灣的森林資源調查記錄資料記載,台灣杉最高可達90公尺。

世界上最高的樹長葉世界爺一赤杉。

澳洲的杏仁按樹林,皆為樹高百公尺以上之巨木,其中最高者高達156公尺,相當於50層的高樓,為現今所發現地球上最高之樹體。
地球上最粗的樹及最古老的樹種
意大利西西裡島上的一裸栗樹,其樹身周長為56公尺,目前被認定為地球上最粗的一棵樹。地球上現存在歷史最久的種子值物為中國的銀杏樹;銀杏樹又名公孫樹,二億年前的石灰紀及二疊紀間即開始繁衍,全盛時期約在一億五千萬年前的侏羅紀迄下白堊紀間的植物。地球演化歷史中,銀杏家族陪同恐龍等古生物一路走來,歷經千錘百鍊繁衍至今,其驚人生命力,成為地球上現存最古老植物,乃有「活化石」之號稱。

意大利西西裡島上的一裸栗樹,其樹身周長為56公尺,目前被認定為地球上最粗的一棵樹。

中國的銀杏樹;銀杏樹又名公孫樹,其驚人生命力,成為地球上現存最古老植物,乃有「活化石」之號稱。
檜木生活習性
紅檜與台灣扁柏具有相似之生態幅度,然因紅檜較台灣扁柏有較強之耐濕性及耐貧瘠性,故常分布於潮濕狹谷之山腹、溪谷,而台灣扁柏則常生長於急峻山腹及嶺線之處。在同一地區,台灣扁柏與紅檜的分布大抵可分為:上坡以台灣扁柏為主的針闊葉林,坡中則為台灣扁柏與紅檜混生,坡下則以紅檜為主。土壤方面,台灣扁柏喜土層較淺、多石、排水良好的乾性灰化土或棕色森林土,紅檜則喜濕性灰壤或棕色森林土。從台灣山地植群垂直海拔分佈梯度來看,檜木林屬礫林帶,其植物組成大致以檜木為主要優勢樹種。紅檜主要生長在海拔1200一2200公尺之間,而台灣扁柏則位於1500一2600公尺之間,大略圍繞山脈山頂成橫向帶狀分布,生長環境終年雲霧繚繞形成雲霧林,多呈檜木純林狀態,向低海拔河谷延伸時僅成少數而與闊葉樹混生;隨著海拔高度增加,檜木逐漸減少,至海拔2800公尺以上時,逐漸轉變為鐵杉與冷杉林之分布。
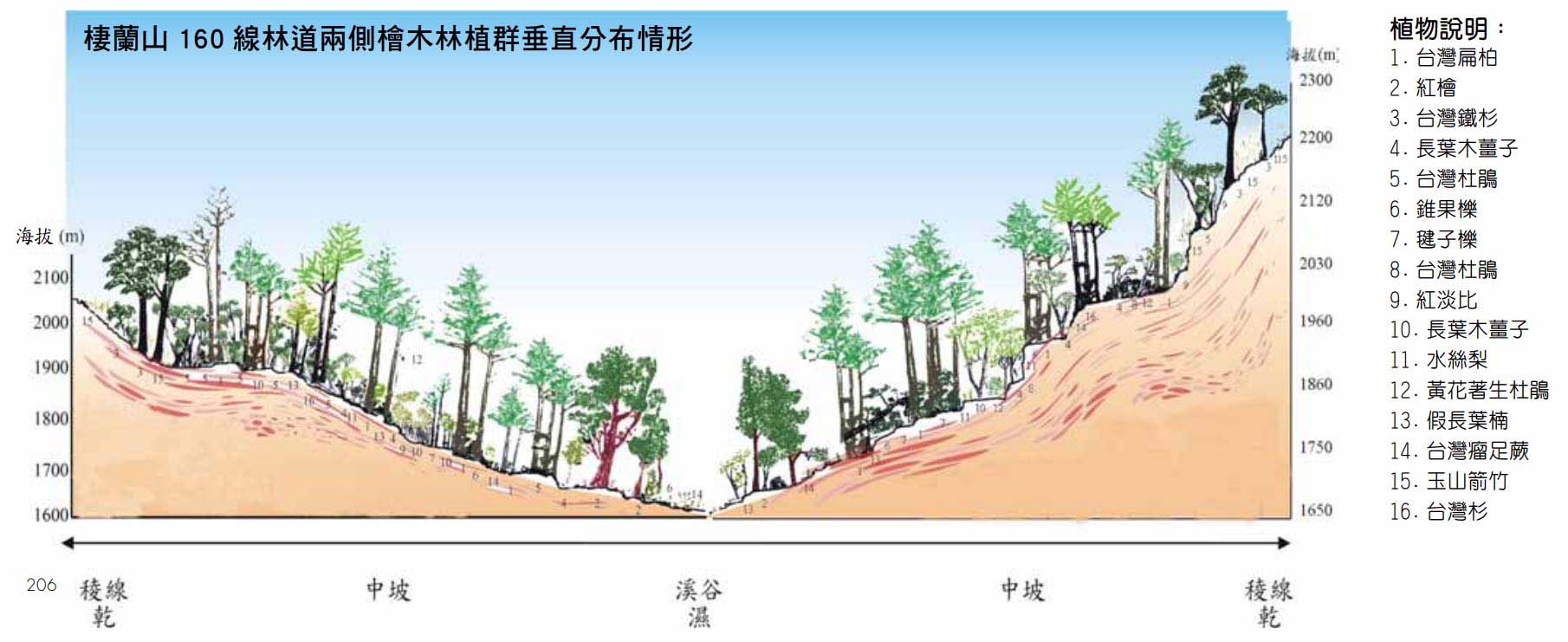
臺灣檜木林垂直分佈情形
由於雲霧林氣候恆定性高,環境適合各種物種生存,形成生物多樣性較高,檜木林區即因位處雲霧林帶,因此林分組成結構亦較為多樣化。由植群多樣性調查發現,影響檜木林區植群變異之環境因子,主要為海拔與地形位置。一般而言,當海拔高度逐漸增加至雲霧帶時,其物種多樣性有增加之趨勢,但海拔高度若繼續上升時,物種多樣性反而下降。檜木林區之物種多樣性組成情況為:低海拔植群社會內物種多樣性高而植群社會間物種多樣性則較低,相對地高海拔植物社會內物種多樣性較低,植群社會間物種多樣性則較高,而海拔差距越大呈現之生態多樣性越大。
檜木為高大長壽之針葉喬木,並為構成檜木林型森林之優勢樹種,在玉山、阿里山、雪山山脈及中央山脈北端等處,均有數量豐富之檜木天然林。由於檜木木材性質強韌、木理優美,利用價值高,為台灣針葉一級木,自日本據台之際即不斷開發伐採,至民國65年台灣林業經營改革方案實施後,始不再伐採天然林。檜木樹齡長壽生長緩慢、木肌密緻,多以巨木形態呈現,由於檜木為淺根性根系,加上高山土壤貧脊土層較淺,易因老死或風害等擾動,造成檜木天然林內,產生相當數量之枯立木及風倒木。檜木木材腐朽分解相當緩慢,倒木長久堆積於地表,有礙其天然下種著土;另由於在林下環境日照不足,及下層闊葉林木、地被植物尤其是密生之玉山箭竹影響,嚴重阻礙檜木之天然下種更新。因此,檜木種苗多出現於林冠孔隙或倒木之上,長期演替下形成林分上層多為老熟檜木,下層稚木數量較少之老熟林相。天然擾動如颱風、崩塌、火燒等,為促進檜木更新之動力;這些天然擾動可造成森林環境空問釋放,有利於下層檜木稚樹行“前生樹更新”(advanceregeneration)或是上層檜木之種子飄落進行“下種更新”(seedingregeneration)。
檜木年輪與生長
面臨太平洋,颱風地震等大然擾動頻繁;當檜木毬果於冬季成熟之際,束北季風造成林區多雨濕潤之氣候形態,有利於種子飄落林地後之發芽更新。台灣扁柏林之伐木根株切口,進行樹輪分析,近千年的樹輪模式顯示該地台灣扁柏更新,主要受到小尺度擾動所形成之單株或塊狀更新以維持當地族群,並不需要大規模的擾動來促進更新。山樹輪生長釋放現象推論台灣扁柏苗可忍受林下遮蔭,直至林冠孔隙形成,再快速生長,以前生樹更新為其更新形式,亦可由空隙形成後行下種更新。
樹輪氣候學,分析樹木年輪以推論氣候變遷的方法。
中緯度的喬木樹幹之橫切面,最容易看見從樹幹中心到樹皮之間具有許多樹輪,顏色深淺交替排列。凡一深一淺樹輪的組合,正好代表一年,稱為年輪。愈往樹幹中心的樹輪形成的時代愈早。分析樹輪橫斷面,可探討紀錄下來的各項環境因子的變化,例如氣候,水文、環境污染,火山噴發、人類和部落遷徙。
樹木年輪中,凡淺色的樹輪代表樹木在春夏季作徑向生長的時期,因樹幹形成層的細胞生長力旺盛,體積大,細胞壁薄,故產生淺色的樹輪,稱為「早材」;反之,到了秋冬季生長力降低,故形成層的細胞壁相對較厚,體積也較小,故產生深色的樹輪,稱為「晚材」。由於早材和晚材交替出現,故層次和界限分明,因此可以判斷出一棵樹的樹齡和區分每一年輪的相對樹齡。若己知樹皮年輪的真實年代,則可以進一步推知該樹誕生的絕對年代。
有時年輪中藏有許多變異樹輪,例如在某一年中的春夏季突然氣候變得不尋常的嚴寒和特別乾旱,則早材可能不會出現,於是產生兩道深色樹輪緊鄰在一起,稱為「雙輪」。雙輪或甚至多道深色樹輪聚在一起,一般稱為「偽輪」。此外可能因整年或許多年天氣長久嚴寒和乾旱,樹木細胞停止了徑向的生長和分裂,因此具有「缺輪」的現象。若忽略雙輪的存在,則在決定樹木的樹齡時,可能少算一年,但如果具有缺輪現象,則至少可能少算一年,甚至更多年,但無法確知到底少算了多少年。
某一地區許多樹木的樹輪樣本,經過統計分析,可以建立起該地區特有之樹輪變化與氣候和環境因子的關聯性,進而可以逆推當地曾經發生過的氣候變化,火山噴發歷史和環境變遷等重大自然事件。早期樹輪氣候學較為考古學界所推崇,但近年也受到全球變遷研究的重視。
臺灣學者針對氣候變遷較為敏感的高山地區採集樹輪加以分析,已推知500年來山區氣候變化形態。進一步對比氣象觀測紀錄,可互相佐證,探討臺灣中長期氣候變化的成因。
檜木形態比較
台灣扁柏與紅檜之形態比較表 |
||
| 項目 | 紅檜 | 台灣扁柏 |
| 樹冠 | 樹冠呈橢圓形 遠視呈淺紅棕色 枝條柔軟略呈下垂 |
樹冠呈圓錐形 遠視呈青黑色 枝與樹幹略呈―直角 |
| 樹幹 | 常有分枝、根張較大 樹皮淺裂 樹皮較薄 年輪生長不均勻 |
通直 樹皮深裂 樹皮較厚 年輪生長均勻 |
| 年輪 | 橫切面早晚材移行緩變而明顯 | 年輪甚狹,早晚材移行急變而明顯 |
| 樹皮 | 樹皮呈淡灰褐至紅褐色 樹皮較薄縱向淺溝裂,又稱 “薄皮仔" |
樹皮呈紅褐至暗褐色 樹皮較厚縱向淺裂或長條剝落,又 稱“厚殼仔" |
| 葉 | 質呈厚色較淺 疏生枝上、覆瓦狀排列 鱗狀葉先端尖銳 葉裡稍現紅色 |
粗壯呈色略深 密生枝上、交互對生 鱗狀葉先端稍鈍 葉裡稍現白色 |
| 毬果 | 形小、橢圓形 果鱗與種子數較多 |
形大、球形 果鱗與種子數較少 |
氣味 |
具有怡人的芳香氣味 |
木材具芳香帶辛辣氣味 |

紅檜一樹幹會分叉樹皮平滑而薄(薄皮)樹冠枝條多上揚

台灣扁柏一樹幹直挺樹皮粗而厚(厚殼)樹冠枝條多下垂

紅檜的樹皮較薄,所以又被稱為「薄皮仔」。

扁柏的樹皮較厚,所以俗稱「厚殼仔」。

紅檜葉

台灣扁柏葉

紅檜果實

台灣扁柏果實

紅繪毬果

台灣扁柏毬果

紅繪苗

台灣扁柏苗
參考文獻:
我見青山多嫵媚
https://www.fcea.gov.tw/uploads/filedownload/00000080.pdf
臺灣檜木之分布與特徵比較
陳盈如 張上鎮
https://www.tfri.gov.tw/main/download.aspx?dlfn=06.105-18_臺灣檜木之分布與特徵比較.pdf
以臺灣檜木著稱的紅檜(Chamaecyparis formosensis Matsumura)與臺灣扁柏(C. obtusa var. formosana Hayata),包裹著臺灣一個世代經濟的成就與溫暖的記憶,至今仍為高雅而優質的象徵,讓人為之著迷。紅檜與臺灣扁柏為臺灣最具代表意義且高經濟價值之樹種,半世紀以來眾多研究人員為此傾注心力深入探討,本文回顧紅檜與臺灣扁柏生育地、生理生態、化學成分、形質特徵與生物活性相關研究之發展以及二者之比較討論。
緒 言
今約4600年前( 約莫中國的新石器時 代),埃及人自黎巴嫩輸入黎巴嫩雪松(Cedrus libani)木材作為建造埃及金字塔之用,這是人 類歷史上第一筆自境外伐取木材之濫觴,亦 是木材供人類締造古文明的具體貢獻。由於針葉樹種木材優美的紋理、獨特的香味與耐久的特性,常被用來建築帝王宮殿及造船之用,爾後的蘇美(Summerians)、亞述(Assyrian)、西 臺(Hittites)、米諾斯(Minoans),以及巴比倫 (Babylonians)等古文明發展過程中,木材皆為重要的建築材料(Farjon 2008),時至今日,木材與人類的生活更具緊密關係。 柏科 (Cupressaceae) 的木材為針葉樹 種中密度與硬度最高之材種,例如紅檜 (Chamaecyparis formosensis)與臺灣扁柏(C. obtusa var. formosana)為臺灣原生的貴重針葉樹,通常合稱為「檜木」。紅檜木理通直、花紋美麗,具耐腐、耐蟻及耐濕性,且有良好的加工性,為國內極貴重的裝潢與家具材料(Hong and Wang 1990);臺灣扁柏則木肌細緻又具濃郁香氣,其價格更高於紅檜(Chiu et al. 1995)。二者在臺灣林業發展中都占有獨特為其他樹種不可及的地位,自日據時期「出口高價檜木購入低廉松杉」的木材貿易政策,乃至臺灣光復後大量外銷檜木以賺取外匯,對臺灣的經濟發展具有重大貢獻。然而也緣於過去之大面積皆伐作業,導致原生針葉林面臨伐盡之危機,直至1989年林務局改為公務預算後方才終止(Hung 1971, Jen 1995, Lee and Hsu 2010)。根據1995 年完成的第3次臺灣森林資源及土地利用調查報告,臺灣檜木天然林面積為48,500公頃、人 工林面積為24,800公頃(Taiwan Forestry Bureau 1995);而2015年完成的第4次全國森林資源調查結果,臺灣檜木天然林面積為29,720公頃、人工林為17,915公頃,檜木之總蓄積量為2518 萬立方公尺(Qiu et al. 2015),二次調查之檜木天然林的面積相差達18,800公頃,其中緣由尚待林務局公佈詳細資料方可得知。
檜木曾支撐臺灣林業的蓬勃發展,從生態演化的功能與林木經濟利用的角度,檜木林 絕對是臺灣珍貴且極需保護的重要自然資產。 檜木生長極為緩慢,紅檜人工林的輪伐期約為 80~120年(Lo-Cho et al. 1987),而臺灣扁柏天然更新林木更需經350~400年的生長才能達到50 cm之胸徑(Lin 1988)。臺灣林業人員不僅致力於臺灣檜木的復育與造林實務工作,同時在育 林、生態、經營與木材利用等相關領域也有廣 泛的研究,發表研究文獻將近200篇。究竟紅檜 與臺灣扁柏在生育地微環境的選擇上有哪些喜好?它們有哪些獨特需求與調適能力?本文將過去50多年來研究人員在紅檜及臺灣扁柏累積 的豐碩研究成果,做一簡單的綜合整理以供關心者共享,也請不吝指正。
分布與生育地
過去曾有學者指出,根據化石分析結果,推測六百萬年前檜木應廣泛分布於世界各地, 因地球氣候與環境之變遷,冰河退卻,導致檜木瀕臨萎縮與滅絕(Liu et al. 2009)。另有學者認為檜木在冰河時期的大遷徙中,由北方逐漸南移而擴大分布範圍,又因陸域板塊的分離與漂移而造就今日的分布(Liu 1966)。目前,檜木存在有5種及1變種,主要分布在北美西海 岸、東海岸、日本以及臺灣等近海岸山脈等局部地區。由於洋流及高速氣流(jet stream)的穿越,這些地區維持相對較為潮濕且溫暖的氣候型態,適於檜木生長(Barbour 2007),為構成北半球太平洋沿岸的山地雲霧森林(mountainous cloud forest)之重要樹種;不同樹種的葉部阻力特徵也使其可適存於不同地區氣候的變異(Liu and Zobel 1980)。因此,有學者推論檜木的分布應是受到水分的限制(Laderman 1998)。這6種檜木即北美西岸的羅生扁柏(C. lawsoniana)、 北美東岸的側葉扁柏(C. thyoides);日本的日本扁柏(C. obtusa)、日本花柏(C. pisifera);臺灣則有紅檜(C. formosensis)及1變種臺灣扁柏(C. obtusa var. formosana)。其中,臺灣為檜木分布於最南端,北美與日本檜木皆生長於暖溫帶針 葉林,僅有臺灣位於亞熱帶。如今,位於棲蘭 山的檜木林不僅是全亞洲僅存的原始檜木林, 更是全世界唯一殘存最大面積的檜木天然純林 (Chi 2008)。文化部曾於2002年將「棲蘭山檜木林」列為臺灣世界遺產潛力點之一(Lei 2014), 期成為全人類共同保護的資產。
雖然紅檜與臺灣扁柏通稱為「檜木」,但 它們的分布環境、生態特性仍有差異。根據林 業研究人員的野外調查與研究,紅檜分布北起插天山稍北處(24°49′N),南至六龜、南大武山一帶(22°35′N);臺灣扁柏則自北部的插天山、鳶嘴山(24°46′N)至南部阿里山、秀姑巒與玉山一帶(23°28′N) (Matsuura 1942, Lin and Kuo 2003)。兩樹種在海拔分布範圍卻有所重疊, 臺灣扁柏分布海拔為1300~2900 m,最旺盛帶為1500~2600 m;紅檜分布海拔比臺灣扁柏稍低,為800~2900 m,最旺盛帶約為1300~2100 m (Chang 1963, Lee 1962)。植群分布調查結果另顯示,不同研究區域所呈現的植物社會與植群型也有所差異,概略而言,臺灣扁柏常見於較高海拔,與紅檜、臺灣鐵杉( Tsuga chinensis var. formosana)、臺灣杉(Taiwania cryptomerioides)、巒大杉(Cunninghamia konishii)等針葉樹形成不同程度之混生或自成純林;而紅檜多見於較低海拔,除與臺灣扁柏、 臺灣杉、鐵杉混生外,亦常與松類及闊葉樹混 生,偶為純林(Chang 1963, Liu 1975b)。
此外,兩樹種之成木與幼苗的分布特性亦有所不同,臺灣扁柏多分布於臺灣北部,紅檜則常見於南部;以中央山脈做為區隔,臺灣扁柏多見於中央山脈西側,紅檜則於東側較 多;以坡向與位置而言,紅檜大多分布於西南南至西北北方向之山坡中、下部及峽谷和溪谷之間,土層為濕潤的灰化土或山地黃棕壤土; 臺灣扁柏則多分布於東北至西南西方向山坡之 上、中部及近嶺線地帶,土壤多為多石礫且排水良好之乾性灰化土或富腐植質的黃壤土、灰棕壤土及黃棕壤土(Liu 1975a, Hung 1971)。在天然更新方面,臺灣扁柏幼苗可在成熟母樹林下生長;紅檜天然更新小苗則較為少見,其幼苗多生長在空曠的生育地,而在成熟林中的幼苗則多見於樹冠鬱閉破裂之正下方或在其外1 m以內的地區(Liu 1975a),可見二者天然更新能力確存有差異。茲將上述2樹種之分布與生育地特性整理如表1。
Table 1. Comparison of distributions and habitat characteristics between Chamaecyparis formosensis and C. obtusa var. formosana
| 特徵 | 紅檜 | 臺灣扁柏 | 參考文獻 |
| 分布海拔 | 800~2900 m | 1300~2900 m | Liu 1961, Lee 1962, Chang 1963 |
| 最盛分布海拔 | 1300~2100 m | 1500~2600 m | Chang 1963 |
| 分布 | 北起24°49′N之北插天山,南至22°35′N之六龜、大武山一帶;多見於中央山脈東側 | 北起24°46′N鳶嘴山 一帶,南至23°28′N 之阿里山、玉山及秀姑巒一帶;多見於 中央山脈西側 | Matsuura 1942, Lin and Kuo 2003 |
| 分布坡面 | 山坡的中、下部,常見於山窪地、潮濕峽谷與溪谷 之間,但偶亦有分布於高海拔上坡之乾溝生育地上 | 多分布於山坡上、中部及近嶺線地帶 | Liu 1975a |
| 習性 | • 較臺灣扁柏耐陽,然幼樹仍須庇蔭。 • 天然更新甚少見,幼樹多為空曠地演替初期之幼林 • 較臺灣扁柏有較強之耐貧瘠及耐濕性。 |
天然更新似無問題,成熟林之幼樹少見於樹冠鬱閉破裂之下方 | Liu 1975a, Ho 1988 |
| 林型 | • 純林,或與扁柏、鐵杉、雲杉或其他針、闊葉樹 混生 • 與臺灣扁柏、臺灣杉、鐵杉、松類及闊葉樹混生 • 多與臺灣扁柏以及溫、寒帶針闊葉樹如松類、鐵 杉、方杉、楠木類、櫧類、臺灣赤楠、槭樹類等 共生,闊葉樹混交林,鮮為純林 • 常和臺灣扁柏及其他針葉樹木混生或形成純林。與假長葉楠、長葉木薑子混生或成純林 |
• 常成純林,但在下緣則與紅檜相混交 • 與臺灣鐵杉、臺灣 杜鵑、巒大杉、臺 灣杉與木荷混生或 成純林 |
Chang 1963, L i u 1 9 7 5 b , Lo-Cho 1987, Ho 1988, Lin 1988, Cheng 2005, Huang et al. 2007 |
| 混交林型 | 紅檜臺灣扁柏、闊葉樹混交林型,分布於海拔高 1000~2200 m | Chang 1963 | |
| 紅檜、臺灣扁柏混交林型,分布於海拔高 1400~2100 m | |||
| 紅檜、臺灣扁柏、鐵杉混交林型,分布於海拔高 1700~2500 m | |||
| 土壤 | 土層厚度中庸至淺薄濕潤地,為濕性灰化土或棕色森林土;土層深厚的濕潤地段,土壤多為灰化土或山地黃棕壤土 | 土壤較淺、多石礫而排水良好之乾性灰化土或棕色森林土;富腐殖質的黃壤、灰棕 壤土及黃棕壤土 | Hung 1 9 7 1 , Liu 1975a, Lin and Kuo 2003, Cheng 2005 |
生理生態特性
早期學者的研究提出了紅檜與臺灣扁柏 族群分布特性及生長環境的差異,而後續的研究分別探討水分與光度等微環境因子對紅檜及臺灣扁柏苗木生理、生長表現之影響,並進一 步分析微環境因子對兩樹種天然更新的限制關係。例如:林讚標等人在1991年對紅檜與臺灣扁柏花粉之萌發條件進行比較,結果顯示紅檜花粉萌發之pH值範圍較臺灣扁柏廣,紅檜花粉 在pH 4~8均可萌發,而臺灣扁柏花粉只在pH 4.5~6.5間方可萌發(Lin et al. 1991)。此外,在模擬的微環境條件下證實,紅檜及臺灣扁柏種子在不同水分條件下的發芽能力亦有所不同, 紅檜種子無論是在淹水或水分短缺的逆境條件下,都比臺灣扁柏種子維持較高的活力與發芽 率,顯示各種水分逆境對臺灣扁柏種子發芽的限制較劇,紅檜種子之發芽對水分逆境之耐受性較強(Lin and Kuo 2003)。至於水分條件對紅檜與臺灣扁柏苗木的生長與生理性狀之影響, 紅檜苗木對水分的需求及蒸發散量顯著大於臺灣扁柏苗木,在水分充足的條件下,紅檜和臺 灣扁柏之光合作用能力無顯著差異,但紅檜苗木之苗高生長量較臺灣扁柏苗木大、具較高之競爭能力;而在低水勢環境下,臺灣扁柏苗木 對水分具有較高的利用效率,能進行光合作用維持苗木生長最低需求;同時,臺灣扁柏脯胺 酸的累積量隨缺水程度的增加而顯著的增加,而紅檜則無,顯示臺灣扁柏具有藉由脯胺酸以適應乾旱的「保護機制」,故而歸納出在水分短缺之微環境中,臺灣扁柏苗木較紅檜的適應 能力更高(Wang 2004)。
在光環境方面,賴宜鈴(2006)在探討光度改變對紅檜與臺灣扁柏小苗生長的研究結果得 知,紅檜與臺灣扁柏在幼苗時期對光度改變具有差異表現。在低光量微環境下,紅檜與臺灣 扁柏小苗之光合作用特性雖無顯著差異,但兩者適應陰暗環境的機制卻有所不同:臺灣扁柏以調整葉部形態的方式、紅檜則以增加莖的長 度來因應;而在高光量環境下,紅檜小苗無論在存活率、光合作用速率、葉相對生長率或生物量都明顯較臺灣扁柏高(Lai 2006)。張桓顥 (2007)的研究亦獲得類似的結果,他發現紅檜苗木對於光資源之競爭能力較強,在不同光度處理下,紅檜之高生長、苗徑與生物量皆明顯 高於臺灣扁柏,且紅檜之暗呼吸速率與光補償點皆低於臺灣扁柏,顯示紅檜在低光環境下較 臺灣扁柏具有競爭優勢(Chang 2007)。
根據上述研究,我們可歸納出紅檜和臺灣扁柏生長表現對微環境需求之差異:在發芽階 段,紅檜較耐水分逆境;在幼苗成長階段,相較於臺灣扁柏,紅檜有較低的光補償點,只需較低光照度便可達到光合作用與呼吸作用的損 益平衡,進而累積有機物,且較低的暗呼吸速率 亦顯示其本身對糖類、澱粉、脂質、蛋白質和有機酸等能量的消耗較低,此種「開源又節流」的 生理特性,是使紅檜無論在高光或低光下皆較臺灣扁柏具較高生長優勢的可能原因之一,這些結果與野外觀察扁柏生長較慢,紅檜生長較快的現 象相符(Hung 1971, Lee 1962),也因而為臺灣扁柏材質較紅檜更為細緻之可能緣由(Lin 1988)。 另值得一提的是,雖然臺灣扁柏花粉或種子之發芽條件皆較嚴苛且易受水分逆境所限制,但幼苗階段對水分逆境的調適能力其實較紅檜幼苗高, 水分利用效率與抵抗乾旱的機制亦優於紅檜幼 苗,這樣的保護機制除了累積脯胺酸外,是否還有其他代謝機制的作用,值得我們深入探討。這些生理生態特性似乎也與天然更新幼苗之存活與 生長表現相呼應,在林地,紅檜的發芽率及死亡率較高,顯示兩樹種的幼苗對微環境適應的差異 (Chung et al. 2011)。若將此與前述「臺灣扁柏幼苗可在其成熟母樹林下天然更新,而紅檜天然更 新則較為少見」之說法做進一步思考,或可推論土壤水分狀況可能為影響兩者更新與分布的重要因子。
無論如何,兩樹種苗木對於不同微環境因子或逆境的差異表現,都可能是影響兩者的逆 境適應性、競爭力,進而導致其在林地自然環境下之生長、更新與分布有所不同的原因,也或許還有更多未知或複合的影響因子有待我們 進一步探討與釐清。茲將上述2樹種之生理生態特性整理如表2。
Table 2. Comparison of physiological ecology characteristics between Chamaecyparis formosensis and C. obtusa var. formosana
| 特徵 | 紅檜 | 臺灣扁柏 | 參考文獻 |
| 淹水逆境 | 可忍受淹水而發芽 | 發芽力減退 | Lin and Kuo 2003 |
| 低水勢逆境 | 於-1.5 MPa之水勢下可維持活力14天而無發芽率下降情形, 超過14天則發芽率減退 | 在7天內發芽率即減退甚多而達顯著差異 | Lin and Kuo 2003 |
| 光合作用 | 光合作用能力潛勢高於紅檜的苗木 | Wang 2004 | |
| 供水充足 | 苗高生長量較臺灣扁柏大 | Wang 2004 | |
| 氣孔導度 | 較大 | Wang 2004 | |
| 水分利用效率 | 較高 | Wang 2004 | |
| 低光下 | 增加莖長度爭取光資源 | 使葉部面積增加以適應陰暗環境 | Lai 2006 |
| 高光下 | 比扁柏具有競爭力 | Lai 2006 | |
| 暗呼吸速 | 較低 | Chang 2007 | |
| 光補償點 | 較低 | Chang 2007 |
形態特徵
前人的研究結果指出,紅檜是在2百萬9千年前和日本花柏分化,而臺灣扁柏約在1百萬3 千年前和日本扁柏分化(Wang et al. 2003),紅檜和臺灣扁柏chloroplast DNA的差異僅有0.57% (Liao et al. 2010, Lei 2014),二者親緣關係頗為緊密,形態外觀也非常相似,但仔細觀察仍可發現許多特徵之差異。例如紅檜的樹形整體枝條較稀疏而下垂、臺灣扁柏的枝條則是密生而平展,紅檜樹皮較薄且縱向溝裂較淺、臺灣扁柏樹皮較厚且溝裂較深,紅檜毬果呈橢圓形、 臺灣扁柏則為球形。若以葉子來區分,3年生以下之紅檜小苗葉子通常仍留有針狀葉,且冬天葉色常呈紅褐色;臺灣扁柏小苗葉子通常不見針狀葉,且終年呈翠綠色,葉背有白蠟;紅檜成熟喬木的葉子呈鱗狀,先端尖銳,逆撫有刺手感;臺灣扁柏成熟木之葉子亦呈鱗狀,但先 端較鈍,逆撫不扎手,葉背有粉白蠟(Lin 1988, Liu et al. 1994, Lai 2006)。
材質特徵
紅檜與臺灣扁柏同為臺灣頂級的針葉樹種木材,皆具有極佳的耐蟲蛀、耐腐朽性與美麗的色澤紋理外觀,然而木材的材質與物理特性 仍有些許差別,也各有不同的喜好者。紅檜與臺灣扁柏的心材率分別為73.88及83.85% (Wu and Hsieh 1988),臺灣扁柏之生材較輕,紅檜 因含水量較多而較重(Hung 1971);紅檜之心邊材區別明顯,材色呈淡紅色,臺灣扁柏之 心邊材區別較不明顯,材色偏黃褐色;紅檜木材有怡人的芳香氣味,臺灣扁柏木材之香氣則較濃郁且帶辛辣氣味(Cheng 2005, Chen et al. 2009)。紅檜木材的整體意象感受是溫暖的,而臺灣扁柏木材則典雅、精緻、溫暖、柔和與自然的感受(Chen et al. 2009)。整體而言,二者皆具有極佳的物理特性,是做為家具、建築與雕刻藝術等之上材,為極珍貴的森林資產。茲將紅檜與臺灣扁柏之葉子與木材之形態、形質特徵整理如表3。
Table 3. Comparison of leaf and wood characteristics between Chamaecyparis formosensis and C. obtusa var. formosana
| 特徵 | 紅檜 | 臺灣扁柏 | 參考文獻 |
| 樹形 | 枝條稀疏而下垂 | Ho 1988, Lin 1988, Liu et al. 1994, Lai 2006 | |
| 樹皮 | • 樹皮呈淡灰褐至紅褐色 • 樹皮較薄縱向淺溝裂,又稱 “薄皮仔" |
• 樹皮呈紅褐至暗褐色 • 樹皮較厚縱向淺裂或長條剝落,又 稱“厚殼仔" |
|
| 葉形態 | 葉成鱗片狀互生,先端尖銳而扎手,3年生以下小苗通常可在苗莖上發現針狀葉遺留 | 葉成鱗片狀對生,先端略鈍形,鱗形葉較薄,小枝上面之葉露出部分菱形,小枝下面之葉被白粉,針狀葉於3至5個月後即脫落。 | |
| 葉色 | 幼葉冬天常呈紅褐色 | 終年呈翠綠色 | |
| 毬果 | • 毬果橢圓形 • 果鱗兩兩相對而生,果鱗 10~13枚 • 每果鱗有種子1~2枚 |
• 毬果球形 • 果鱗8~10枚,中央有一小突起 • 每果鱗具種子2~5枚 |
|
| 花粉 | 平均長寬軸30.7×31.8 µm | 平均長寬軸33.8×35.1 µm | Lin et al. 1991 |
| 木材 | • 邊心材區別稍明顯 • 巨木常會因蓮根菌腐蝕而樹幹中空 |
• 邊心材境界不明顯 • 樹幹不易空心 |
Hung 1971, Cheng 2005, Chen et al. 2009 |
| 年輪 | 橫切面早晚材移行緩變而明顯 | 年輪甚狹,早晚材移行急變而明顯 | |
| 材質 | 木肌細緻、木理通直 | 木肌細緻、通直均勻 | |
| 色澤 | 材色呈淡紅色 | 材色偏黃褐色 | |
| 整體意象感受 | 溫暖的 | 典雅、精緻、溫暖、柔和與自然的 | |
| 氣味 | 具有怡人的芳香氣味 | 木材具芳香帶辛辣氣味 | |
| 生材種量 | 因含水量較多而較重 | 生材重量較輕 | Hung 1971 |
| 細胞 | 樹脂細胞豐富,多在秋材部分 散狀分布或成切線狀;縱向薄 壁細胞橫向壁節狀加厚較明 顯,且木質線薄壁細胞細胞橫 向壁具紋孔。 | 樹脂細胞多切線狀分布於春材部分 | Chen et al. 2009, Li and Wang 2013 |
化學成分特徵
有關臺灣扁柏精油成分的研究,Kafuku (1931)的研究中曾提及最早乃起始於Uchida (1916)利用乾餾法(dry distillation)萃取臺灣 扁柏木材之精油,發現其精油含有d-α-pinene 及l-cadinene等成分(Kafuku et al. 1931)。接著Tsuchihashi與Tasaki (1920)改以水蒸氣蒸餾法(steam distillation)獲得d-α-pinene、d-αterpineol、isoborneol、d-cadinene等 類化 合物及dicyclic sesquiterpene alcohol與酸類 (acid)、酚類(phenolic)等成分。爾後1926~1963 年間,Hirao、Kafuku、Nozoe及林耀堂等人亦曾分別對臺灣扁柏木材精油成分展開一系列深入的研究(Kafuku et al. 1931, Lin et al. 1955a, b, 1942, 1963, Lin and Wang 1960);其中,Nozoe於1938年自臺灣扁柏心材精油中分離出一個烯醇類化合物(enolic compound),期間歷經第二次世界大戰延宕,直至1948~1949年 才確定其結構為2-hydroxy-4-isopropyl-2,4,6- cycloheptatrienone,也就是現今大家所熟知的具特殊七圓環的tropolone-扁柏醇(hinokitiol) (Fang and Cheng 1992),因其結構特殊及極佳的生物活性而備受關注。
臺灣扁柏葉子之精油成分則於1931年由 Kafuku等人開始研究,在較低沸點的部分獲得 fatty acid (C10H16O2)、hinokic acid (C16H24O2)、 d -sabinene 、d - α -pinene 、p -cymene 、α-terpinene、γ-terpinene和α-thujene等成分,以及一個新發現的單環化合物(chamene) (Kafuku et al. 1931);沸點較高的部分則發現了 2個tricyclic sesquiterpene成分以及d-cadinene 等化合物(Kafuku and Nozoe 1931)。他們也分析紅檜葉子精油之組成,這也是紅檜精油成分最早之分析記錄,紅檜葉子精油主要以 l-α-pinene最多,另包括單 類(l-camphene、 dipentene、α-terpinene, γ-terpinene、cineol和 borneol以及倍半 類(cadinene、humulene),以及一個沸點較高的倍半 類化合物cadinol (Kafuku and Ichikawa 1931)。他們後續的研 究則進行紅檜材部精油的成分分析,結果顯示 紅檜材部精油除了葉子所具有的d-α-pinene、 d-camphene及α-terpineol等單萜類化合物之 外,還富有l-cadinol、allyl-protocatechin 等,同時亦發現了4種新的萜類成分(benihiol (C10H18O)、benihinol (C10H16O)、benihinal (C10H14O)和d-sesquibenihene (C15H24) (Kafuku and Ichikawa 1933)。
二次戰後,臺灣早期的科學研究人員篳路藍縷,自簡易的研究設備獲得得來不易的成 果,1960年代王光燦首次以薄層層析技術(thin layer chromatography)分離紅檜葉子之抽出成分,使酚類化合物之分離效果獲得改善(Lin et al. 1958, Wang 1959, Fang and Cheng 1992),此後陸續進行紅檜葉子(Lin et al. 1962)、根部 (Nozoe and Toda 1966)、材部(Asao et al. 1968, Cheng et al. 1971)及皮部(Fang et al. 1986b)等抽出成分之分析,獲得許多過去難以分析的微量成分。1986年,方俊民等人首次利用氣相層析儀進行紅檜葉子精油之成分分離,共鑑定出 41個化合物(Fang et al. 1986a),不僅解析度提升,且可獲得各個成分所占之相對比例。自此植物化學的研究蓬勃發展,紅檜與臺灣扁柏各部位之精油、抽出成分以及揮發性化合物都累積了許多的研究成果,例如鄭玉瑕、郭悅雄與鍾婷婷等教授之實驗室皆曾對紅檜與臺灣扁柏之葉、毬果、木材甚至樹皮之抽出成分進行深 入的分析,臺灣扁柏各部位之抽出成分皆以雙 類化合物為主,其次為倍半類化合物;而 紅檜之葉、樹皮與毬果抽出成分主要仍為雙萜類化合物,另包括有類黃酮類、木酚素與芳香族等化合物,木材抽出成分則以木酚素類及類木酚素類為主(Xu 1982, Zhou 1990, Xu 1993, Lin et al. 1999, Chen 2000, Chan 2004, Chien 2004, Ko 2005, Wu 2005, Chen 2006, Kuo 2007, Chen 2008, Chen et al. 2008)。
整體而言,由近年的研究結果得知,紅 檜材部之精油以myrtenol (13.53~48.89%)、 myrtanol (12.69~13.17 % )、δ-cadinene (26.3%)及α-eudesmol (12.90~18.06%)等成分之相對含量最高(Kuo 2004, Wang et al. 2005, Kuo et al. 2007, Chen et al. 2015),而 α-pinene(57.32~85.97%)則是紅檜葉子精油最主要的成分(Fang et al. 1986a, Su et al. 2006, Chen et al. 2011)。關於臺灣扁柏材部精油的成分研究,除最具代表性的hinokitiol之外,鄭玉 瑕教授實驗室亦曾透過柱層分析、氣相色層分 析、薄層分析與再結晶等方法解析出精油成分 包括α-pinene, α-copaene, β-elemene等共13種化合物(Chen 1975)。至於近年臺灣扁柏葉子精油成分的相關研究,Su等人於2006年之研究報告指出,臺灣扁柏與紅檜葉子精油皆同樣以 α-pinene為其主要成分,且含量皆>70% (Su et al. 2006)。然而,Chen等人為更進一步確認 臺灣扁柏屬植物葉子精油成分之差異,於臺灣 北、中、南、東等地共採取43棵單株、超過130 個樣本進行其成分分析,其結果卻顛覆了過去 的認知,一般被認為普遍大量存在針葉樹精油中的基本成分α-pinene,在所分析的臺灣扁柏葉 子精油樣本中含量都非常低(0.31~1.41%),主要成分反而是β-elemol (24.33%)、thujopsene (widdrene) (18.43~34.84%)以及cis-thujopsenal (25.12%)等倍半萜類化合物,同時根據不同地區來源與成分特徵形成了三種化學品系;不僅如此,含有豐富的thujopsene更是臺灣扁柏重要的化學特徵,在全球其他扁柏屬樹種中,僅日本扁柏(C. obtusa, 0.15~3.6%) (Park et al. 2003, Hong et al. 2004, Yang et al. 2007)、日本花柏 (C. pisifera, 0.95%) (Zhang et al. 2009)及已自扁柏屬除名的拿加遜扁柏(C. nootkatensis, trace) (Cheng and von Rudloff 1970)含有此成分,但含量亦非常低。目前,紅檜葉子中α-pinene生合成酵素之基因已被成功選殖(Chu et al. 2009), 關於臺灣扁柏類生合成酵素之研究仍缺如, 因此,究竟臺灣扁柏具有何種神秘的特質,使得它不同於一般針葉樹,僅含有極微量的 α-pinene,卻又有別於扁柏屬之其他樹種,獨具有豐富的thujopsene,著實是一個非常值得我們 探討掘挖的「彩蛋」,深入研究後或許將有意想不到的驚喜收穫!茲將這些較具代表性之二次代謝物成分特徵整理如表4。
Table 4. Comparison of secondary metabolites between Chamaecyparis formosensis and C. obtusa var. formosana
| 種類 | 部位 | 紅檜 | 臺灣扁柏 | 參考文獻 |
| 精油 | 葉子 | α-pinene為主要成分 | thujopsene, β-elemol, cisthujopsenal等最具特色 | Fang et al. 1986a, Su et al. 2006, Chen et al. 2011 |
| 木材 | myrtenol, myrtanol, δ-cadinene及α-eudesmol等 分相對含量最高 | hinokitiol, α-pinene, α-copaene, β-elemene等 | Lin et al. 1955a, Chen 1975, Kuo 2004, Wang et al. 2005, Kuo et al. 2007, Chen et al. 2015, | |
| 抽出物 | 葉子 | 雙萜類化合物為主,其次 為倍半萜類化合物 | 雙萜類化合物為主,其次 為倍半萜類化合物 | |
| 木材 | 以木酚素類及類木酚素類為主 | 雙萜類化合物為主,其次為倍半萜類化合物 | ||
| 樹皮 | 雙萜類化合物為主 | 雙萜類、固醇類等化合物 | Zhou 1990, Ko 2005, Wu 2005, Kuo 2007 | |
| 毬果 | 雙萜類, 倍半萜類, 木酚素類等化合物 | 雙萜類, 倍半萜類, 木酚素類等化合物 | Xu 1993 (PhD thesis) | |
| 揮發成分(SPME, HS) | 葉子 | germacrene D, α-pinene相對含量最高 | thujopsene, compound A, sabinene, thujopsenal相對含量最高 | Lin et al. 2011, Chen et al. 2015 |
| 木材 | myrtenol為主要成分 | Wang et al. 2006 | ||
| 芬多精 | 葉子 | α-pinene, β-Myrcene為主 | Lin et al. 2015 |
二次代謝物之生物活性
二次代謝物通常是指有別於植物初級代謝 物(如氨基酸、核苷酸、多糖…等)之外的次級 代謝產物,每一種植物在特定的環境條件下,具有生成特定化學成分的特性,而為該種植物的生理生化特徵,也形同演化成一個獨特的化學工廠,在生長的嚴酷世界裡製造出求生的物 質。過去曾認為這些物質是不具重要功能的無 用產物,然而近年來的研究結果發現,某些特定的二次代謝物可能存在於某些特定植物中並 賦予生態上的特殊功能,在眾多物種中,植物歷經天擇的考驗,可能蘊藏著具有改善人類生活或解決人類重要醫藥問題的契機。
眾所周知,臺灣檜木具有極佳的抗白蟻、耐腐特性,早期的研究認為,木材精油含油量之 多寡可能是影響其抗白蟻性質之原因,紅檜、臺灣扁柏、臺灣肖楠以及香杉等含油量較高之木材都具有優良之抗白蟻性質(Wang et al. 1987);紅檜木材與根部的chamecynone是賦予良好抗白蟻能力的重要成分(Harayama et al. 1977, Fang et al. 1986b),而臺灣扁柏葉子精油也已證實有良好的抗白蟻活性(Cheng et al. 2007)。此外,紅檜木材精油還具有優良的抗病媒蚊幼蟲與衣魚幼蟲 能力,可望做為環保殺蟲劑之用(Kuo 2004, Kuo et al. 2007),其材部之抽出成分亦被證實具有抗發炎活性(Hsieh et al. 2007),而臺灣扁柏心材抽出成分則具有抑制癌症細胞生長之潛力(Chien et al. 2007),其皮部抽出物與葉子熱水抽出物則具有優良的抗氧化活性,尤其棲蘭山地區臺灣扁 柏葉子熱水抽出物不僅收率高、抗氧化活性優良且毒性低,深具開發潛力(Marimuthu et al. 2008, Cheng et al. 2014)。
最早由臺灣扁柏木材分離出而命名的hinokitiol乃是柏科木材特有之成分,為木材抽出成分中最著名的抗細菌成分之一,許 多研究證實紅檜與臺灣扁柏心材精油能抑制金黃色葡萄菌(Staphylococus aureus)、產氣 性桿菌(Enterobacter aerogenes)與肺炎桿菌 (Klebsiella pneumoniae)等細菌之生長(Wu and Wang 1990),而紅檜木材精油還具有抑制褐腐菌(Laetiporus sulphureus)及白腐菌(Trametes versicolor)等真菌生長的效果(Wang et al. 2005),賦予木材優良的耐腐朽性。另一方面,檜木精油散發的香氣不僅迷人,許多研究結果證實其對人體自律神經總活性有提升作用(Chen et al. 2015),並具有舒壓、解憂以及減輕疲勞 等功效(Hsieh et al. 2001),更讓社會大眾對其在芳香療法上之應用趨之若鶩。茲將紅檜與臺灣扁柏之二次代謝物成分活性整理如表5。
Table 5. Bioactivities of secondary metabolites from Chamaecyparis formosensis and C. obtusa var. formosana
| 種類 | 部位 | 紅檜 | 臺灣扁柏 | 參考文獻 |
| 精油 | 葉子 | 抗白蟻活性(anti-termitic activities) | Cheng et al. 2007 | |
| 木材 | 抗病媒蚊幼蟲(antimosquito larval activity)、衣魚毒殺效果 (anti-silverfish activity) | Kuo 2004, Hsieh et al. 2007 | ||
| 抗細菌(antibacterial activity) | 抗細菌(antibacterial activity) | Wu and Wang 1990 | ||
| 殺蟲活性(insecticidal activity) | Kuo et al. 2007 | |||
| 抗蟎活性(antimite activity) | Oribe and Miyazaki 1997 | |||
| 抗真菌活性(antifungal activity) | Wang et al. 2005 | |||
| 枝條 | 抗病原菌活性(antipathogenic activities) | Ho et al. 2012 | ||
| 抽出物 | 葉子 | 抗氧化活性(antioxidant activity) | Cheng et al. 2014 | |
| 木材 | 抗真菌活性(antifungal activity), 抗發炎活性(anti-inflammatory) | 抑制癌症細胞生長之潛力(potent cytotoxic activity against cancer cells) | Kuo 2004, Chien et al. 2007, Hsieh et al. 2007 |
結語
無論「棲蘭山檜木林」最終是否被列為世界遺產,紅檜與臺灣扁柏就歷史文化、生物保 育、自然教育意義與生物科學研究來說,都是臺灣極為珍貴的資產。過去林業機構曾努力復育造林,然而現今研究人員更應積極思索如何透過科學手段固守並延續這些珍貴的樹種,例如透過建立有效的種子來源評估系統,維持種原之多樣性並避免不良之遺傳變異,同時建立妥善完整的育苗與造林策略,並評估對整體生態系的影響(Huang et al. 2007),如此才能真正達到維持檜木林生態系多樣性,並回復其生態 健康、完整性與永續生存之目標。此外,面對全球大環境迅速而劇烈變化的衝擊,檜木林的 命運又該何去何從?研究人員曾利用氣溫因子的溫量指數透過統計方法建立情境模擬,推估當全球溫度上升,檜木林潛在分布將大幅減少 (Chiu and Huang 2008)。然而植物面對環境的變化會有各種適應或抵抗逆境的生存機制,檜 木的生存分布究竟還受哪些因子所調控?其適應環境的機制為何?這些仍有待我們未來做更深入的研究探討。