大元山植物生態之研究
── 章樂民 1961‧10 ──
Ecological Studies On the Vegetation of Mt.Ta·Yuan by Lo-Min Chang
資料來源:林業試驗所報告第070號
https://www.tfri.gov.tw/main/other_in.aspx?mnuid=5381&modid=1&cid=13&nid=193
![]()
![]()
![]()
![]()
![]()
一、緒言(Introduction)
植物生熊之調查與研究,為林學之基礎科學,森林施業上諸如立地因子,樹種選定,造林,撫育,更新,砍伐等各作業,均須依據植物生態學之原理,以解決森林造成之適當手段,大元山事業區為本省光復後國人新開闢之林區,具有熱暖溫三帶之森林,植物豐富,林相優美。本研究於民國 四十七年十月及四十八年十一月,先後前往元山兩次作實地調查與研究,所得資料經攜回進行整理與分析,爰草此報告以供營林上之參老焉。
本計劃之完成得國家長期發展科學委員會之補助,主作期間蒙業師林所長渭訪之指導與校閱, 蘭陽林區管理處大元山工作站予以工作上之便利及協助,謹此一併誌謝。
二、目的及方法(objeotive and method)
本計劃之目的在初究大元山植物生態全貌,探詞環境因子對植物生態之影響,各森林帶植物社會之組成,分佈,生態演進之研究,指示植物之探求,籍供該事業區今後營林上之參考。茲述其方 決如下:
1·環境因子(Environment factors)
(1)氣候(Climate):蒐集大元山山區之山地氣象資料供研究參考用。
(2)土壤(Soils):調在每一標準區(Sample plots)土壤種類,割面各層次之深度, 色澤,溼潤度,母岩種類,含石量等。
2.概況調查:就大元山事業區內作全面踏查,以明瞭該區植物分佈狀態,生態概觀,並採集植物標本等,然後設置標準區。
3.標準區之設置:海拔高300m以下因多農墾地故未設有標準區外,海拔高300m以上依垂直分佈分南向與北向,按不同林況每升高海拔50m,設置標準區一個共計64個。標準區面 積為10×10立方米,每一標準區記載土壤剖面形態,環境,立木度,傾斜度,海拔高度,植物種類之高度,直徑,株數,生活形等,個體頻度則根據全部事業區植被分佈概況估計並以簡單符號表示之:優勢d.(Dominant),多數.(Abundant),少數f.(Frequent)點生o.(Occasional),群生l.(Local),稀r.(Rare),甚稀v.r.(Very rare)
4.植物生態之研究:依標準區調查結果,進行下列研究:
(1)植物區系(FIora):大元山植物種類及分佈狀態。
(2)植物生活形(Lifefol'ms):採用Raunkiaer氏生活形分類式,分析統計其結果。
(3)植物社會分類(Plant cormmunity):確定大元山植物群系(Formation),群叢(Associntion),類別名稱,明瞭安定植生與不安定植生,並進行下列各項統計分析:
1.頻度(Frequency %)%某樹種佔據標準區數目,對標準區總數比,以百分率表示之即(某樹種佔據標準區數/標準區總數)X100
2.密度(Density)某樹種總株數,對標準區總數之比即(某樹種總株數/標準區總數
3.相對密度(Relative density %)某樹種株數對標準區內各樹種總數株和之比,以百分率表示之即(某樹種總株數/標準區內各樹種總株數) ×100
4.優勢度(Dominance) 某樹種胸高斷面積和對標準區總數之比,以百分比率表示之,即(某樹種胸高斷面積和/標準區總數)
5.相對優勢度(Dominance%)標準區內某樹種斷面積總和,對各樹種斷面積總和之比,以百分比率表示之,即(某樹種斷面積總和/標準區內各樹種斷面積總和)×100
(4)大元山植物社會組成(Structure),層次(layers),生態演進(Plant succession)等之研究。
三、事業區之概述(Description of the Area)
1.地況(Topography)
大元山事業區,位於本省宜蘭縣境內,東北東向,約在東徑120度10分至121度44分,北緯24度20分至41度41分,向東為羅東事業區,西與太平山事業區毗鄰,南與南澳事業區為界限,向北與羅東事業區交界,事業區之山脈屬於中央山脈系統內,海拔高自300公尺至2,000公尺止,其主要者有大元山1,475公尺,三星山2,265公尺,十六份山1,821公尺,飯包尖山1,680公尺,各山脈又連接貫通,故事業區內之地勢,多屬高峻臉阻,其他無名高山亦復不少,此等山脈貫通於大元山事業區16個林班內,成一馬蹄形盆地,向北北東出口,河流主要以大南澳溪,源出於中央山脈,橫流於事業區內,向北流為大南澳北溪,向南流則為大南澳南溪,東流入海,由於溪流錯雜繁多,地形分割益形險峻,各溪流之源頭基岩露出者頗多,土地崩壞者亦復不少,如大南澳北溪141及142林班之大斷岩,至於局部崩山斷岩多於溪畔見之,一般言之傾斜度均大,惟稜線邊緣部份傾斜度常較緩和故有優美林相之形成。
植物之分佈(Distribution),遷移(Migration),聚集(Aggregation),土著(Ecesis)競爭(Competition),反作用(Reaction),安定(Stabilization)等各種現象告受立地因子直接與問接之影響,其中以氣候與土壤影響最大,此為植物學家公認之事實,過去多將植物認作生長在固定環境中之個體來研究,而缺少對植物社會與環境因子間之認識,庶不知植物社會與環境因子間,乃互為因果,二者關係至切。
2.氣候(Climate)
根據大元山測候站(設於海拔高800m),本所臨時測候站(設於海拔高1,300m),及與大元山交界同緯度之太平山測候站(設州每找高1,900m)測得民國46,47,48年三年來之平均記錄如下:
(1)溫度 (temperatures)
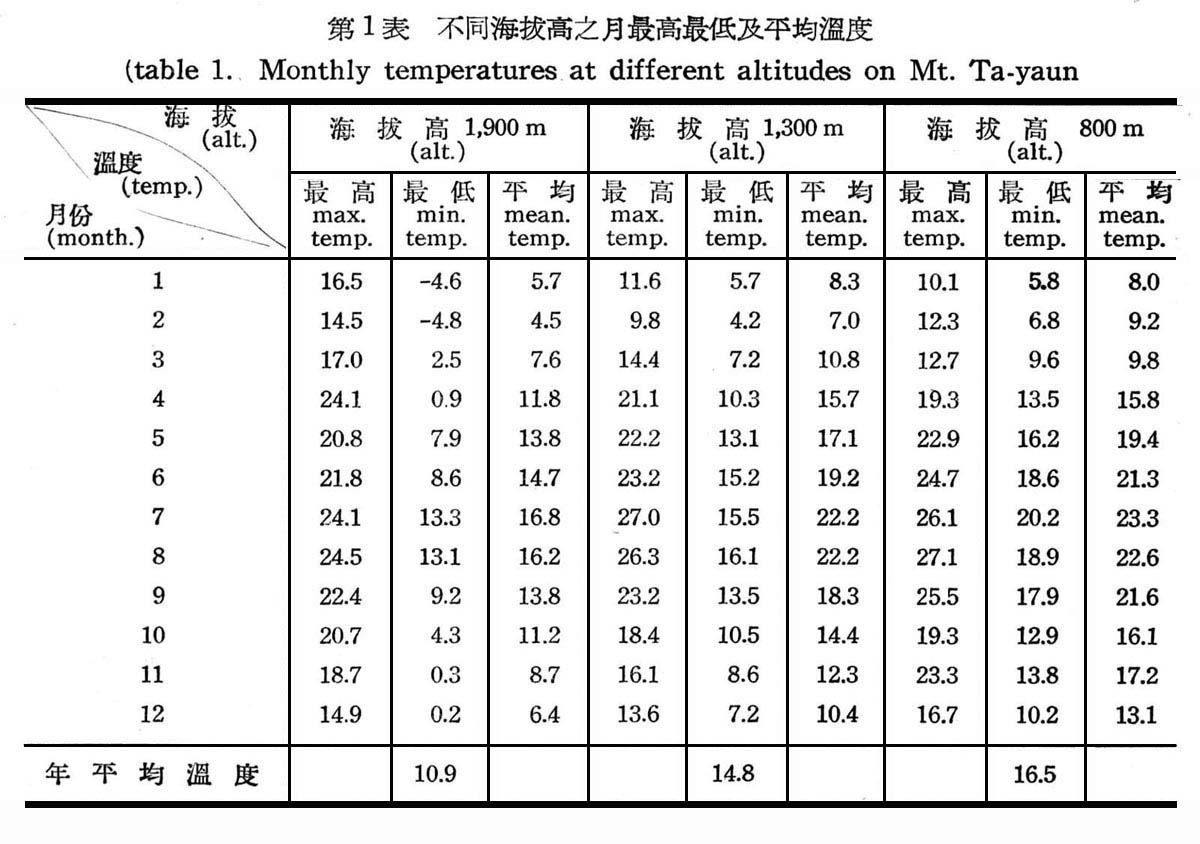
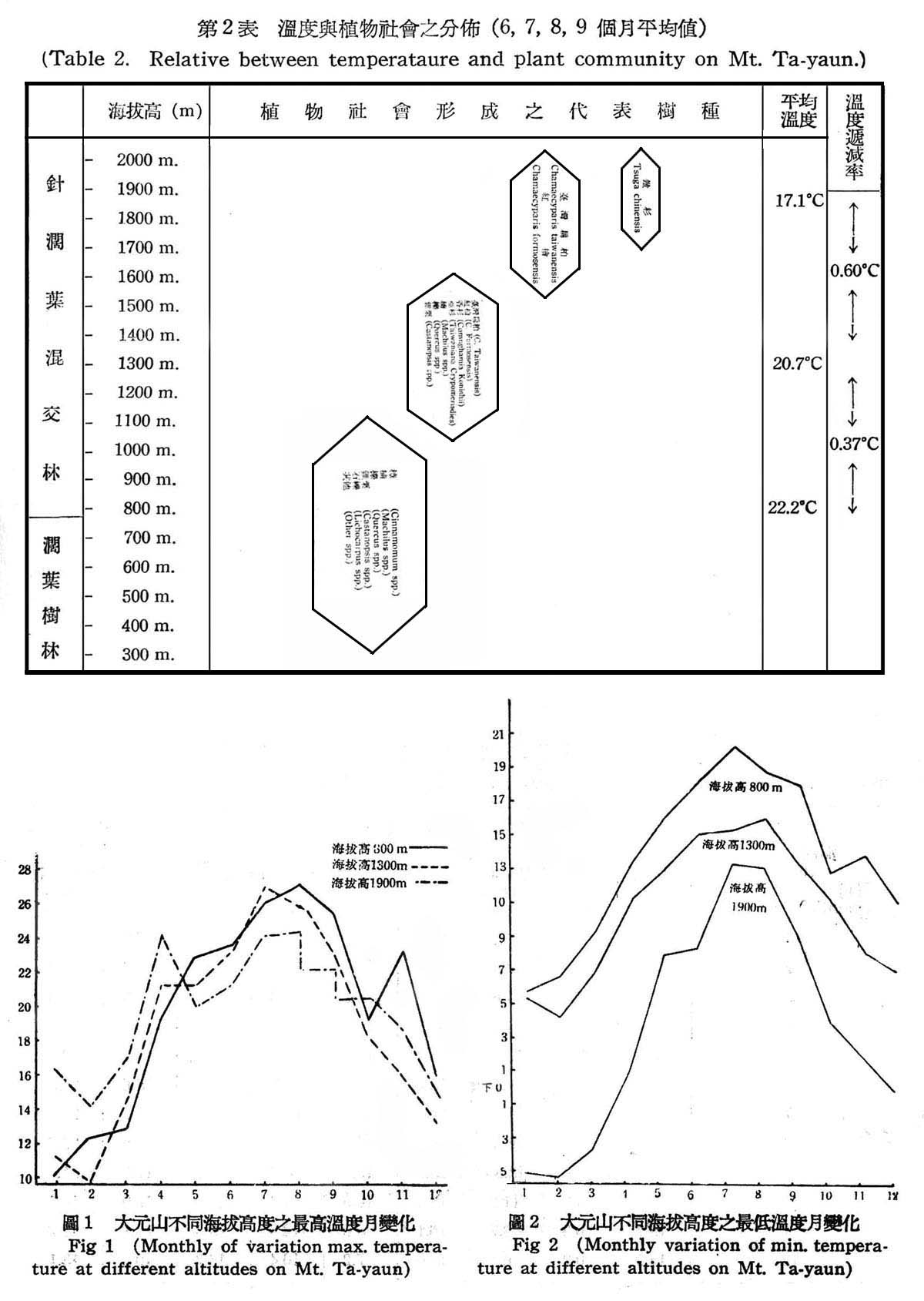
溫度為植物生育必需之條件,植物之種類與分佈,以及植物社會之延續,皆受溫度所支配,溫度之記載有最高溫度,最低溫度,平均溫度等,尤其最高溫度與最低溫度較為重耍,(1)由表2所示,溫度遞減率隨海拔高之上升而降低,植物社會之變化亦隨溫度而急變,植物生長,一年中大致以6,7,8三個月最為旺盛,9月以後則逐漸緩慢,故此四個月內,應為植物生長最適宜季節,以此四個月之平均值,則海拔高800m至1,300m,平均溫度為攝氏22.2度至20.7度每升高海拔100m,溫度垂直遞減率為攝氏0.37度,其所代表樹種為常綠葉樹(evengreen broad leaved tree)趨向針葉樹(Coniferous)海拔高1,300~1,900m,平均溫度為攝氏20.7度至17.1度,每升高海拔100m,溫度垂直遞減率為攝氏0.60度,其所代表樹種趨向於針葉樹純林,至於300~800m雖缺少記錄,但為常綠潤葉樹林殆無疑義。(2)由表1所示,大元山之年平均溫度亦隨海拔高而不同,海拔高800m上下為攝氏16.5度,海拔高1300m為攝氏14.8度,海拔高1900m為攝氏10度,依Engler氏對世界植物種領分佈之解析“植物分佈受溫度之影響,所表現植物各有不同,年平均溫度在攝氏15度以上,20度以下,加以多量濕度,所表現之植物社會為常綠潤葉樹林,年平均溫度在攝氏14度以下,與多量濕度,即為溫帶植物”,頗為符合,大元山終年有充足濕度,海拔高800~1,300m,潤葉樹林生長旺盛,應屬於常綠闊葉樹林帶,海拔高1,300~1,900m,應屬於溫帶針闊混交林,(3)植物能適應一大距離之溫差,即生長各有最適溫度,故其範圍較狹,反之生存所需溫度其範圍較廣,即植物能耐最高與最低溫度程度如何,端視植物種類而異,依表1圖1,2,所示大元山海拔高800~1,300m常綠闊葉樹林帶,最高溫度為八月份攝氏27.l度,最低溫度為二月份攝氏4.2度,海拔高1,300~1,900m為針闊葉樹林帶,最高溫度為八月份攝氏24.1度,最低溫度為二月份零下攝氏4.8度,故二者所需溫度顯然有甚大幅度,故闊葉樹生育時問較長所需溫度亦較高,而針葉樹每生長於寒冷地區,生長時問短,所需溫度亦低。
(2)濕度降雨量降雨日數(Relative humidity,Rainfall, and NumberofRaindays.)
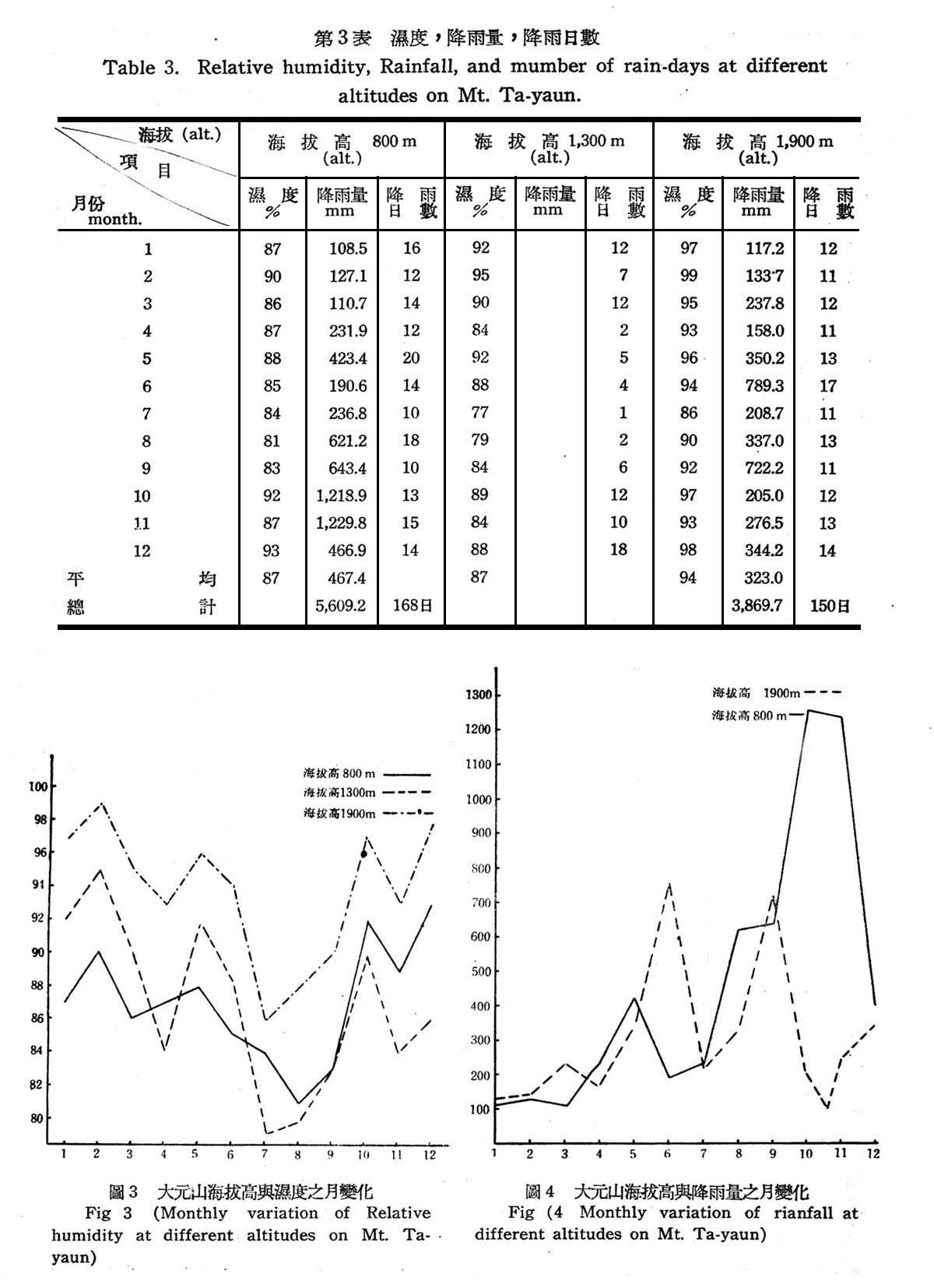
濕度為決定植物蒸發最主耍因子,空氣中水分以蒸發狀態存在,植物在一地區生長全視植物蒸發所失水分,是否超過所吸收者,經常保持70~80%濕度,對植物生長有利,濕度過少則不足供植物之蒸發,濕度過剩則阻礙開花結實,並且易罹病蟲害,一般言之,針葉樹所需濕度較闊葉樹高,針闊葉混交樹林則視混交林木百分率以決定濕度之增減。由表3及圖3,4風示,大元山濕度隨海拔高度而增加,尤其9~12月份濕度普偏上升,海拔高800m常綠闊葉樹林帶,及海拔高1,300m針闊葉混交林帶,年平均濕度均在87%,海拔高1,800~2,000m針葉樹林帶,年平均濕度為94%。故天元山全年濕度甚為平均,非但濕度充足,且巳過剩,由圖15所示,林內苔蘚滿掛於樹幹上即可證明,至海拔高1,800~2,00Om針闊葉混交林帶以針葉樹占百分比較高,因濕度高故適於針葉樹生長,闊葉樹生長則不良,此顯示過剩濕度對闊葉樹乃有不良之反作用。
水分為植物生育必需之物質,植物從土壤中吸收水分,而土壤中水分主視降雨量而決定,一年中降雨量均勻分佈,對植物生長較有利,且降雨量與蒸發量至少需保持平衡狀態,則植物方可充分生育,對植物社會之延續關係密切,由第3表所示大元山海拔高800m常綠闊葉樹林帶,年總雨量為5,609.2m.m,月平均雨量為467.4m.m.,降雨日數全年168日,海拔高1,900m針葉樹混交林帶,年總雨量為3,869.7m.m,月平均雨量為323.0m.m,降雨日數為94日,故一年中雨量尚未見不足,年蒸發量雖缺少資料可供參老,唯根據陳正祥氏調查臺灣山地年蒸發量,大概約在1,000m.m.左右,而大元山為重濕氣候,年降雨量在,869.7~5,609.2m.m.降雨量大於蒸發量甚多,故土壤無缺水現象,植物生長不受阻礙。
(3)風,霜,雪(Wind, Frost and Snow)
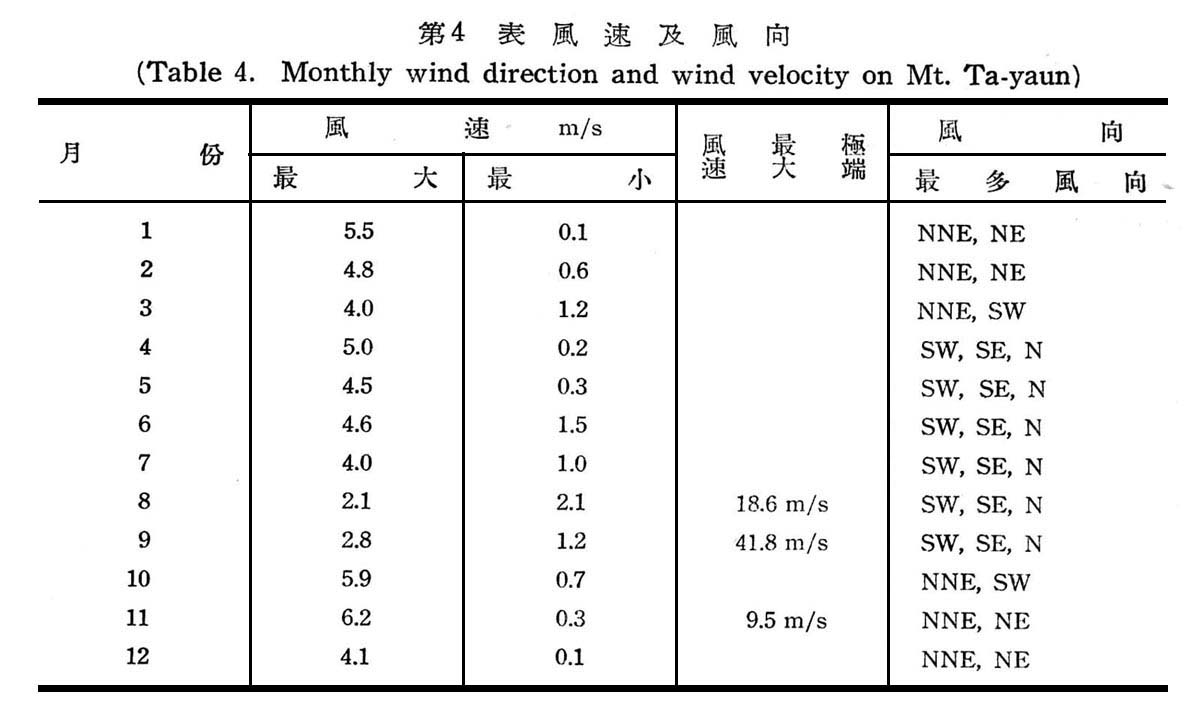
山區之風速與風向,受局部地勢之影響,大元山林區地形為一馬蹄形盆地,受風之影響各有利弊,一般言之,風對植物屆顛分佈,遷移,土著,等均屬有利,大元山紅檜扁柏分佈豐富,殆亦有關係,但如終年均受風襲,林內蒸發加速,土壤乾燥易引起植物生理上之乾燥,強烈東北季風及夏季颱風,則易發生風倒現象,尤其林木生長及幼林木受害最大,如表4所示,大元山一年中,冬季東北季風開始於11月至翌年4月,平均最大風速為5.lm/s,最大風速為11月6.2m/s,極端最大風速亦為11月達9.5m/s,最小風速12月及1月為0.lm/s,夏季則為東南或西南風,惟風力遠不及冬季之東北季風強烈,夏季之季節風,開始於5月至10月,平均最大風速為3.3/s,最大風速為十月5.9m/s,但在此期間一年中受數度颱風之侵襲,其極端最大風速達18.6~41.8 m/s,為害最大,大元山有豐富之降雨量,土壤無缺水現象,植物不至於受生理上乾燥之害,至於冬季東北季風甚少有達風速15m/s以上者方可破壞林木,影響不大,但夏季颱風對林木為害最大,尤針葉樹為害最烈,闊葉樹影嚮皎小。
大元山每年均有降霜與下雪,多發生於12月至2月間,日數不多,維持時間亦短,林地內霜雪,使土壤發生凍結現象,對林木根部有害,針葉樹稚苗受害最易,唯大體上對林木影響不大。
3.地質與土壤(Geology and Soils)
構成大元山森林土壤之母岩為水成岩之粘板岩及頁岩,溪流沿岸則多第四紀洪積層山麓則常見風化極為顯著之崩壞地,粘板岩及頁岩由於地質脆弱,風化容易而成粘質壤土(Clay loam)或粘土(Clay)其分佈範圍甚廣,由海拔高300~1,900m均有,溪流洪積層乃由各種石岩風化沖積而成之沖積土,其有效成份不及粘質壤土及粘土,各種土壤均夾雜有石礫約30~60%不等,海拔高1,800~2,000m,因受氣候因子之影饗,因而生成灰壤(Podsol)即所謂定域土(Zonol Soil),唯其分觸龜圍較狹,一般言之,地勢險峻者,土層較能,傾斜度稍緩者,則土層較深,由於濕度大,多呈濕潤狀態。
代表土壤剖面之形態
(1)沖積土
環境:海拔高720m,天然闊葉樹林之第二期次生林,近河床為沖積小平臺,含石礫量約在60%以上,有效深度不及1m。
I.枯枝落葉未完全腐爛,深度約3cm,呈濕潤狀態。
II.含粗腐植質,深度約1cm,呈濕潤狀。
III.含少量腐植質,富角礫為砂質壞土,暗褐色,深度約4cm呈濕潤狀。
IV.相當於A1富角礫砂質壤土,褐色,深度約30cm
V.母岩:結晶片岩
(2)崩積土
環境:海拔高850m,傾斜度約20度,天然潤葉樹林之第二期森林含石量約在60%,並有大塊頁岩雜於土中,有效深度不及1m.過去為崩壞地。
I.枯枝落葉未完全腐爛,深度約2cm,呈撫渭伏。
II.層次不明,含少量粗腐質,深度約10cm,灰黑色,結構鬆,呈濕潤狀。
III.粘質壤土具有粘性,灰黃色,結構稍鬆,呈濕潤狀。
IV.深度約25cm,雜有角礫,黃色呈濕潤狀,結構稍緻密具有粘性。
V.心土層深度10cm,為風化未完全之母岩,大量雜在粘土中。
Vl.母岩:頁岩
(3)殘積土
環境:海拔高1,000m.近稜線,天然闊葉樹林。有效深度在lm以上。
A00 落葉枯枝未腐爛,深度約3cm。
A0-A1 層次不甚明顯,含粗腐植質,深度約6cm,暗黑色。
A2 含少量腐植質,深度約50cm,雜有細礫,結構稍鬆,略有粘狀,褐色,呈 濕潤狀。
B2 深度約40cm,雜有細礫,結構緻密具有重結性,黃色呈濕潤狀。
C 乃風化未完全之母岩,觸之呈剝落狀,深度約10cm。
D.母岩:粘板岩
(4)灰化土
環境:海拔高1500m,上腹部,上層為天然針葉樹林,下曆為玉山箭竹。
A00 未完全腐爛之枯枝落葉,深度約2cm
A0-A1 含粗腐植質,深度約5cm,結構鬆,黑色呈濕潤狀。
A2含少量腐植質,深度約12cm,略有粘性,褐色呈濕潤狀。
B:深度約9cm,紅黃色,結構稍級密具有粘性之植土,呈濕潤狀。
B:層次不明顯,具角礫,深度約8cm,紅黃色,結構甚為緻密之粘土,呈濕潤狀。
C.風化未完全之母岩,深度約4cm
D.母岩:粘板岩
(5)灰壤(Podosol)
此係受氣候因子之影嚮,而產生之土壤,即所謂定域土(Zono Soil),寒泠多濕之林地,針葉樹林之枯枝落葉分解緩慢且不完全,多量粗腐植質堆積結果而成,此類土壤多分佈於大元山高山寒冷地區,海拔高1,500~2,000m處,上層林木為針葉樹林,下層為玉山箭竹及羊齒類,其土壤斷面形態如下:
A00未腐爛之落葉枯枝深度不足1cm。
A0含粗腐植土,深度約9~15cm,結構疏鬆尤如海棉狀,黑色呈濕潤狀。
A1含多量腐植質,深度約1~7cm,結構稍疏鬆,黑色乃至褐色呈濕潤狀。
A2深度約3~15cm,灰色或灰黑色,結構緻密具有粘性,呈濕潤狀。
B 深度約1~10cm,結構緻密,透水性不良,赤褐色。
C 黃褐色,深度約5~10cm。
D.母岩:粘板岩
綜上所述,大元山之森林土壤,一般言之傾斜大,土層淺,反之傾斜度小者,土層略深,闊葉樹林林地之土壤水分均呈濕潤狀態,通透性良好,多屬酸性,針葉樹林之林地,土壤水份與前者相近唯通透稍劣,多屬強酸性。林地之土壤,其土層之深厚與物理性優良與否,較之有效養分含量多寡,尤為重要。
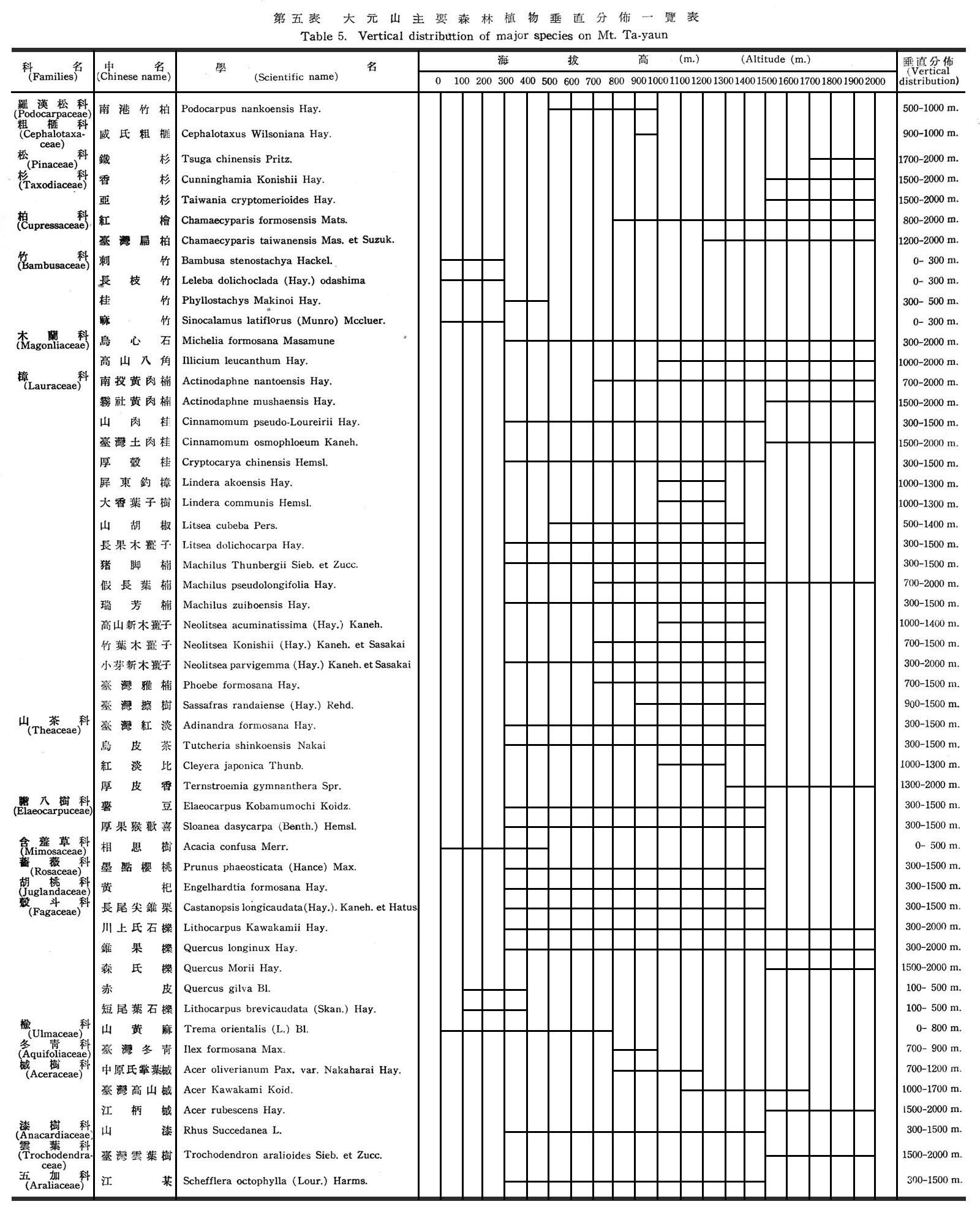
四、大元山植物區系之位置及森林植物垂直分佈概況(Flora and vertical distribution of foresttrees)
關於本島植物區系之區分,各植物學家多有不同之申論,根據工藤博士謂臺灣植物區系,可分為北部小區,中部小區,東部小區,恆春半島小區,蘭嶼與綠島小區,山本由松縛士將全島劃分為大屯山小區,中央山脈小區(西部南部,恆專半島及東部斜面),臺東山脈小區,此外澎湖,蘭嶼綠島各成一小區,李惠林博士主張將全島劃分為本島及恆春二區,劉棠瑞先生則將本島劃分為北部,西部,東部,恆春半島及高地(3,000m以上之地區)五小區,上述諸學者主要皆依植物種類分佈,再依地域而區分,皆有事實之根據。
佐佐木舜一博士認為臺灣全島應列入東亞溫暖區,而將其劃分高地帶小區,及平地小區,後再區分為北部臺灣,臺灣內陸及熱帶海流帶三亞小區,此一區分方法,似略帶有生態之意義,作者認為植物區系之區主要固依種類為重耍,但對於環境因子(氣候土壤等)亦須作考慮蓋環境因子往往影響植物種類分佈,則本島植物小區系之區分,應劃分為高地帶小區,與低地帶小區,前者再區分為北部亞區,中部亞區,東南部亞區,後者再區分為平地亞區,與熱帶海流亞區(包括鵝鑾鼻半島)而澎湖蘭嶼綠島自成一小區,則大元山海拔高500m以下應屬於平地亞區,500m以上應屬於北部亞區範圍內。
大元山森林植物垂直分佈,依本省森林帶垂直分佈言,海拔最高為2,000m,不過在溫帶林之邊緣,猶未超過進入寒帶之針葉樹純林範圍,本島南北緯度不同,森林垂直帶區分略有差異,茲就大元山森林植物分佈現狀,按海拔高度敘述大元山森林植物分佈如下:
1.熱帶林(海拔高100~500m)
本帶森林植物在300m以下者,由於人口稠密,受人為因子破壞,固多農墾地,天然林多成散生或點生狀態,人工林則以相思樹(Acacia confuse Merr.)及樟樹(Cinnamomum Camphora Sieb.)有小面積之純林,麻竹(Sinocalamus latiflorus (Munre) Mcclure)刺竹(Bambusa stenostachya Hackel.),長枚竹(Leba dolichoclada(Hay.)odashima)等均有少數栽培於村落內,海拔高300~500m多為綿延之丘陵地帶,除相思樹及桂竹(Phyllostachya Makinoi Hay.)成塊狀之純林外,其主要森林植物具有經濟價值者有:
長尾尖葉錐栗(Castanopsis longicaudata (Hay•)Kaneh•etHatus.
錐果櫟(Qucrcus longinux Hay.)
短尾葉石櫟(Lithocarpus brevicandata (Skan.)Hay.)
赤皮(Quercus gilva Bl.)
瑞芳楠(Machilus zuihoensis Hay.)
豬腳楠(Machilus Thunbergii Sieb.et Zucc.)
山肉桂(Cinnamomum pseudo-Loureirii Hay.)
長果木薑子(Litsea dolichocarpa Hay.)
厚殼桂(Cryptocarya chinensis Hemsl.)
黃杞(Engelhardtia formosana Hay.)
烏心石(Michelia formosana Mas.)
墨點櫻桃(Prunus phaeosticta (Hance) Max.)
江某(Schefflera octophylla (Lour•)Harms.)
烏皮茶(Tutcheria shinkoensis Nakai)
紅淡(Adlnandra formosana Hay.)
上述森林植物胸高直徑均在15cm以下,最大直徑亦未超過20cm,個體密度分佈亦少,林木經濟價值低微,此外尚有經濟價值更低之小喬木主要者計有:
俄氏虎皮楠(Daphniphyllum Oldhami (Hemsl.) Rosentll.)
紅皮(Styrax suberfolius Hook. et Arn.)
薯豆(Elaeocarpus Kobanumochi Koidz.)
福氏面頭粿(Glochidion Fortunei Hance)
山漆(Rhus succedanea Linn.)
樹杞(Ardisia Sieboldii Miq.)
赤楊葉(Alniphyllum Fortunei Makino)
山柿(Diospyros eriantlla Chaip.)
烏材柿(Diospyros morrisiana Hance)
山黃麻(Trema orientalis (L.) BI.)
野桑(Morus australis Poir.)
山龍眼(Helicia formosana Hemsl.)
山香圓(Turpinia formosana Nakai)
九芎(Lagerstroemia subcosata Koehne)
白桕(Sapinm discolor Muell-Arg.)
野桐(Mallotus japonicas Muell-Arg.)
白匏子(Mallotus paniculatus (Lam.) Muell-Arg.)
臺灣沙朴(Celtis formosana Hay.)
水京金(Wendlandia formooana Cowan.)
稜果榕(Ficus spetica Burm.)
榕樹(Ficus retusa Linn.)
九丁榕(Ficus nervosa Heyne)
楓香(Liquidambar formosana Hance)
構樹(Broussonetia papyrifera (L.) Hertier)
水管心(Saurauja Oldhami Hemsl.)
朱實九節木(Psychotria rubra Poir.)
咬人狗(Laportea pterostigma Wedd.)
金氏榕(Ficus Kingiana Hemsl.)
牛乳樹(Ficus Harfandj Benth.)
蘭盆筆筒樹(Alsophila pustulosa Christ.)
筆筒樹(Cyathea taiwaniana Nakai.)
2.暖帶林(海拔高500~1,500m)
本林帶在700m以下林相無重大變化,主要森林植物仍以熱帶林之森林植物為主體,700m以上林相微有變化,上述陽性樹種,如福氏面頭粿,山漆,紅皮,赤楊葉,俄氏虎皮楠,山黃麻,野桐,白匏仔,水京金及桑科植物,已逐漸減少,繼起者則有:
枇杷葉山礬(Symplocos eriobotryaefolia Hay.)
竹葉木薑子(Neolitsea Konishii (Hay.) Kaneh. Et Sasaki)
臺灣雅楠(Phoebe formosana Hay.)
南投黃肉楠(Actiondaphne nantoensis Hay.)
中原氏掌葉槭(Acer Oliverianum Pax. var. Nakaharai Hay.)
田代氏石班木(Rhaphiolepis indica Lindl. var. Taohiroi Hay.)
臺灣冬青(Ilex formosana Hay.)
小芽新木薑子(Neolitsea parvigemma (Hay.) Kanell. Et Sasaki)
長果木薑子(Litsea dolicliocarpa Hay.)
山櫻桃(Prunus campanulata Max.)
850m以上純然屬於暖帶林之常綠闊葉樹,而熱帶性之森林植物幾已絕跡,此時則有紅檜(Chamaecy Parisformosensis Mats.)之出現,而與在臺灣其他區域出現之高度比較,應屬於最低者,海拔高1000m~1300m以上屬於針葉樹則有臺灣扁柏(Chamaecyparis taiwanensis Mas. Et Suzuk.)南港竹柏(Podocarpus nankoensis Hay.)紅檜等三種,屬于闊葉樹者則有:
1.與海拔高850m以上共通數種:
長尾葉錐栗,錐果栗,短尾葉石櫟,赤皮,瑞芳楠,豬腳楠,山肉桂,長果木薑子,厚殼桂,黃杞,鳥心石,墨點櫻桃,江某,烏皮茶,紅淡,樹杞,赤楊,薯豆,枇杷葉山礬,竹葉木薑子,臺灣雅楠,南投黃肉楠,中原氏掌葉槭,臺灣冬青,小芽木薑子,九芎等。
2.繼起之樹種則有
厚果猴歡喜(Sloanea dasycarpa Hemsl.)
臺灣土肉桂(Cinnamomum osmophloeum Kaneh.)
川上氏石櫟(Lithocarpus Kawakamii Hay.)
澀梨(Malus formosana Kawak.)
大香葉子樹(Lindera communis Hemsl.)
紅淡比(Cleyera japonica Thunb.)
屏東釣樟(Lindera akoensis Hay.)
山胡椒(Litsea cubeba Pers.)
高山新木薑子(Neolitsea acuminatissima (Hay.) Kanch.)
梭葉木薑子Neolitsea acuto-trinervia (Hay.) Kaneh.)
臺灣高山槭(Acer Kawakamii Koid.)
臺灣擦樹(Sassafras randaiense (Hay.) Rehder)
紅柄槭(Acer rubescens Hay.)
銳葉柃(Eurya acuminata DC.)
能高山茶(Camellia nokoensis Hay.)
細葉茶(Camellia tenuifolia (Hay.) Makino)
筆羅子(Meliosma rigida Sieb. Et Zucc.)
威氏粗榧(Cephalotaxus Wilsoniana Hay.)
臺灣山礬(Symplocos formosana Brand)
大葉山礬(衍mplocos theophrastaefolia Sieb. Et Zucc.)
本林帶之闊葉樹較熱帶闊葉樹生長可稱良好,胸高直徑最大者可達50cm以上,平均在20cm以上,林業經濟價值稍高,海拔1,300~1,500m以上闊葉樹林已逐漸開始生長不良,繼起者之優勢種則為針葉樹林。
3.溫帶林(1,500~2,000m)
本林帶依臺灣森林帶論應屬於溫帶林,而未達寒帶林之針葉樹林之純林,而大元山1,500m以上針葉樹占絕對優勢,幾為針葉樹純林,而闊葉樹生長惡劣形成極明之第二層樹冠,主要針葉樹除紅檜(Chamecyparis formosensis Mats.),臺灣扁柏(Chamaecyparis taiwanensis Mas. Et Suzuk.)占最大面積,胸高直徑平均在70cm以上,其他針葉樹則有鐵衫(Tsuga chinensis Pritz.)及少數香衫(Cunninghamia Konishii Hay.),亞衫(Taiwania cryptomerioides Hay.) 等,常綠闊葉樹林多成小喬木及灌木狀,直徑在在15cm以下,主要者則有:
錐果櫟(Quercus longinux Hay.)
高山八角(Illicinm leucanthum Hay.)
厚皮香(Ternstroemia gymnanthera Spr.)
銳葉柃(Eurya acuminate Dc.)
細葉柃(Eurya crenatifolia Yam.)
臺灣杜鵑(Rhododendron formosanum Hemsl.)
森氏杜鵑(Rhododendron Morii Hay.)
臺灣雲葉(Trochodendron aralioides Sieb. Et Zucc.)
豬腳楠(Machilus Thunbergii Sieb. Et Zucc.)
南投黃肉楠(Actinodaphne nantoensis Hay.)
烏心石(Michelia formosana Mas.)
假長葉楠(Machilus pseud01ongifolia Hay.)
山肉桂(Cinnamomum pseudo-Loureirii Hay.)
墨點櫻桃(Prunus phaeosticta (Hance.) Max.)
臺灣上肉桂(Cinnamomum osmophloeum Kaneh.)
川上氏石櫟(Lithocapus Kawakamii Hay.)
短穗花山礬(Symplocos divaricativena Hay.)
枇杷葉山礬(Symplocos eriobotryaefolia Hay.)
小西氏山礬(Symplocos Konishii Hay.)
森氏櫟(Quercus Morii Hay.)
霧社黃肉楠(Actinodaphne mushaensis Hay.)
茲將大元山具有經濟價之主要森林植物垂直分佈,列一簡表如下:
五、植物社會之研究(Study in Plant Communities)
植物社會乃一複雜有限之有機體,具有一定之發育過程與組織,受環境因子所控制,乃環境因子支配下之產物,植物社會最大單位為植物群系(Plant Formation),同一群系主耍以氣候因子與植物生活形(Life Forms)(指優勢種與次優勢種而言)皆田類似下而區分為若干群叢(Plant Association),群叢之下因土壤關係又分為若干植物帶(Zonation)即喬木下分為灌木,草本,著生植物,苔蘚,地衣等,此種縱向之劃分構成多層次(Layer)之樹冠,交互出現,第一層樹冠之疏密,可決定第二、三層之植被種類,頗為重耍,植物社會之區分各有不同之分類形式,大元山之植物社會依作者意見,應稱為常綠木本植物社會,雖然各林帶氣候土壤及植物種類略有差異,但其植物生活形與形態上之構造則大致相若,其區分如下:

1.亞熱帶降雨林群系(Subtropical rain forest formation)
本島熱帶降雨林,僅限於本島南部之恆春半島,即楓港溪,阿塱衛溪以南之地區,四季暖熱,年雨量平均在3,000mm以上,乾燥期甚短,植物生育旺盛,代表此種降雨林主要植物除龍腦香科(Dipterocarpaceae)無分佈外,主耍者以棕梠科(Plamae),大戟科(Euphorbiaceae),桑科(Moraceae)蕁麻科(Urticaceae)等植物,其他地區熱帶降雨林之發育甚為貧弱,所見者均應屬於亞熱帶降雨林,其分佈僅限於海拔高500m以下,除常見之熱帶植物外,並參與若干常綠闊葉樹林帶之樟科(Lauraceae)及殼斗科植物(Fagaceae),實際上可稱為常綠闊葉樹林延長林帶,故其分佈範圍甚狹,本省低海拔地常由於人口稠密,開墾,火災,伐採等現象,甚為普遍,亞熱帶降雨林已難保持其真正熱帶雨林之植物景色。茲就植物群叢(Associatfon),組成層次(layers),胸高直徑與株數分配,天然更新(Reproduction),生態演進(Suecession)等分述如下:
(I)植物群叢概述(Simple description of plant association)
構成大元山亞熱帶降雨林之主要植物草叢(Plant Association)分述如下:
山黃麻─榕中途群叢(Trema orientalis-FicusSpp., Associes)分佈於低海拔100~700m,土壤略乾燥,寫照充足之開墾地或伐採跡地,為第二期森林之早期發 生之植物,在山腳部占最多數,廣汎分佈於全島。
白匏仔中途單叢(Mallotuspaniculatus,Consocies)
分佈於海拔高100~8,000m,土壤乾燥或濕潤均適於生長,平地山麓以至於中海拔之開墾地或伐採跡地佔多數,當其他中性闊葉樹侵入後成被壓木時,則逐漸淘汰,為第二期森林最習見之陽性植物。
桫欏群叢(Cyathea-taiwaniana-Alsophila pustulosa, Association)
分佈於海拔高300~800m,北向蔭濕環境中之常綠闊葉樹林下部,成大面積散生狀態,尤其溪流峽谷最為繁茂,為亞熱降雨林特有之木本羊齒,普遍分佈於全島。
木苧蔴中途團集(Boehmeria densiflora, Colony)
分佈於海拔高100~500m,向陽乾燥環境,溪傍河邊崩壤地或伐採跡地,屢屢成群生狀態,普遍分佈於全島。
山香圓─山龍眼群叢(Turpiniaformosana-Heliciaformosana, Association)
分佈海拔高300~700m,濕潤闊葉樹林中,為亞熱帶降雨林第二層樹冠,一般高度均在4m以下,為亞熱帶降雨林主要宭叢之一。
長尾尖錐栗─黃杞群叢(Castanopsis longicaudata-Engelhardtiaformosana,
Association)
分佈於海拔高300~700m,土壤深厚而潤濕環境中,構成亞熱帶降雨林第一層樹冠,亦為主要群叢之,中性植物社會,為氣候極盛相(climatic climax)
芭西木群叢(Barthea formosana, Association)
分佈於海拔高500~700m,土壤稍濕潤,亞熱帶降雨林鬱閉略破壞,陽光折射林內之林下灌木,散生或成團集狀態,其分佈範圍不若常綠闊葉樹林內之普遍。
火炭母藤中途團集(Polyonum chinense, Colony)
分佈於海拔高300~500m,向陽略乾燥環境中,或鬱閉破壞後林下略有發現,尤其林綠路傍最多,成塊狀分佈。
菅草中途群叢(Miscanthus spp. Associes)
分佈於海拔高400~1,800m,土壤乾燥貧瘠環境中,開墾地,火燒跡地,崩壞地最多,林緣林隙亦有發生,屢屢成大面積分佈,為全省林地習見群叢之一。
芒箕骨中途群叢(Dicranopteris linearis, Consocies)
分佈於海拔高300~500m,向陽乾燥土壤貧瘠環境中局部出現,多生於酸性之紅土,路傍林緣特多。
(2)組成曆次(Structure layers)
構成大元山亞熱帶降雨林組成層次,可分為喬木層(Trees layer),灌木層(Shrubs layer),草本層(Herb layer),及著生植物苔蘚,地衣等,18個樣區測定結果,茲列各表以示之。

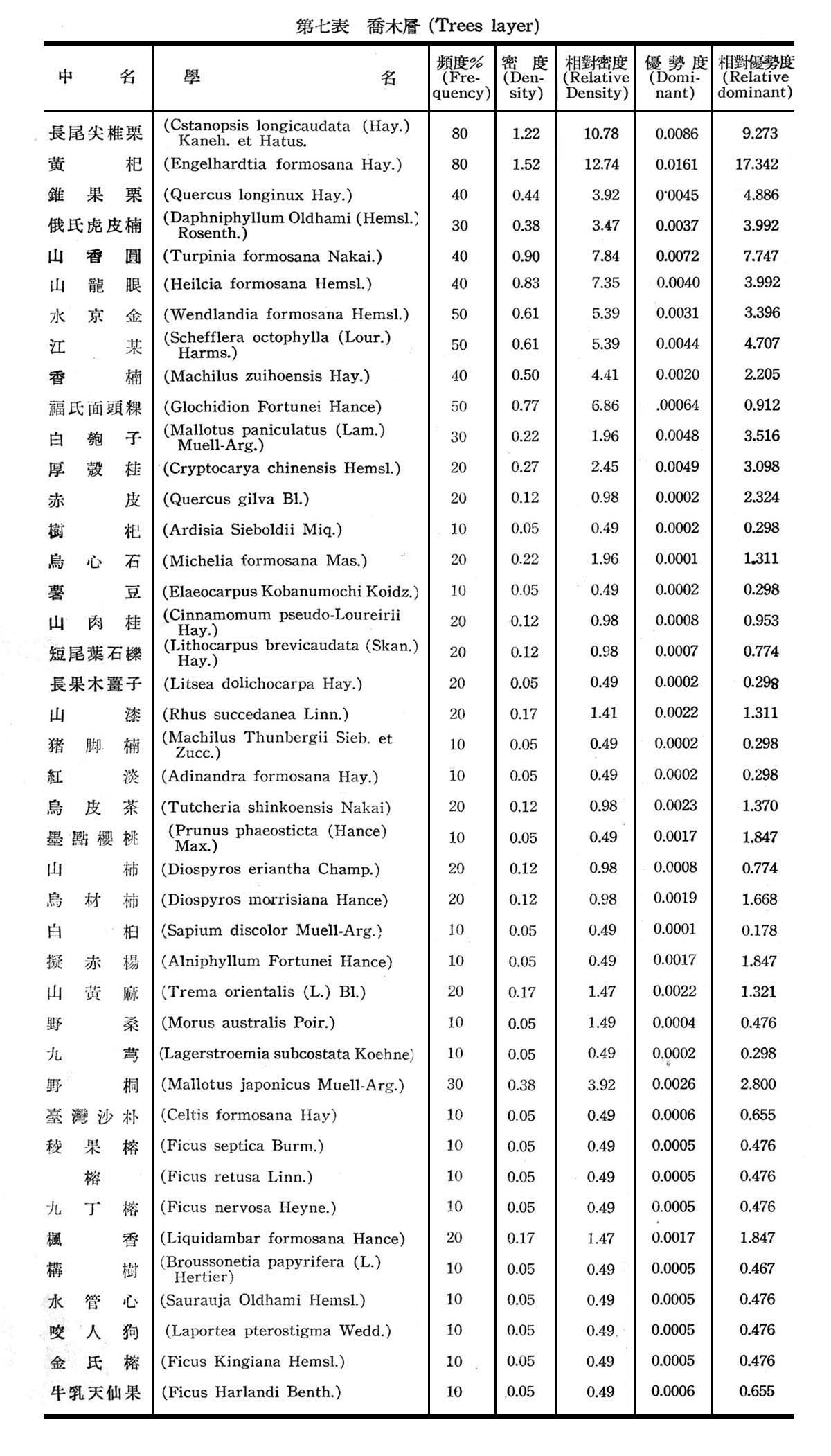


觀上列第七─九表構成亞熱帶降雨林計喬木層42種,灌木層31種,草本層21種,著生植物(羊齒植物)16種,苔蘚植物一種,地衣一種,由表7所示頻度,密度,優勢度均以長尾尖錐栗及黃杞為最大,為優勢種(Dominant Species),錐果櫟,江某,俄氏虎皮楠,山龍眼,山香圓,水京金,厚殼桂,福氏面頭粿,白匏仔等次之,為次優勢種(Co-dominant Species),故構成大元山亞熱帶降雨林,仍以長尾尖錐栗─黃杞群叢為主體。
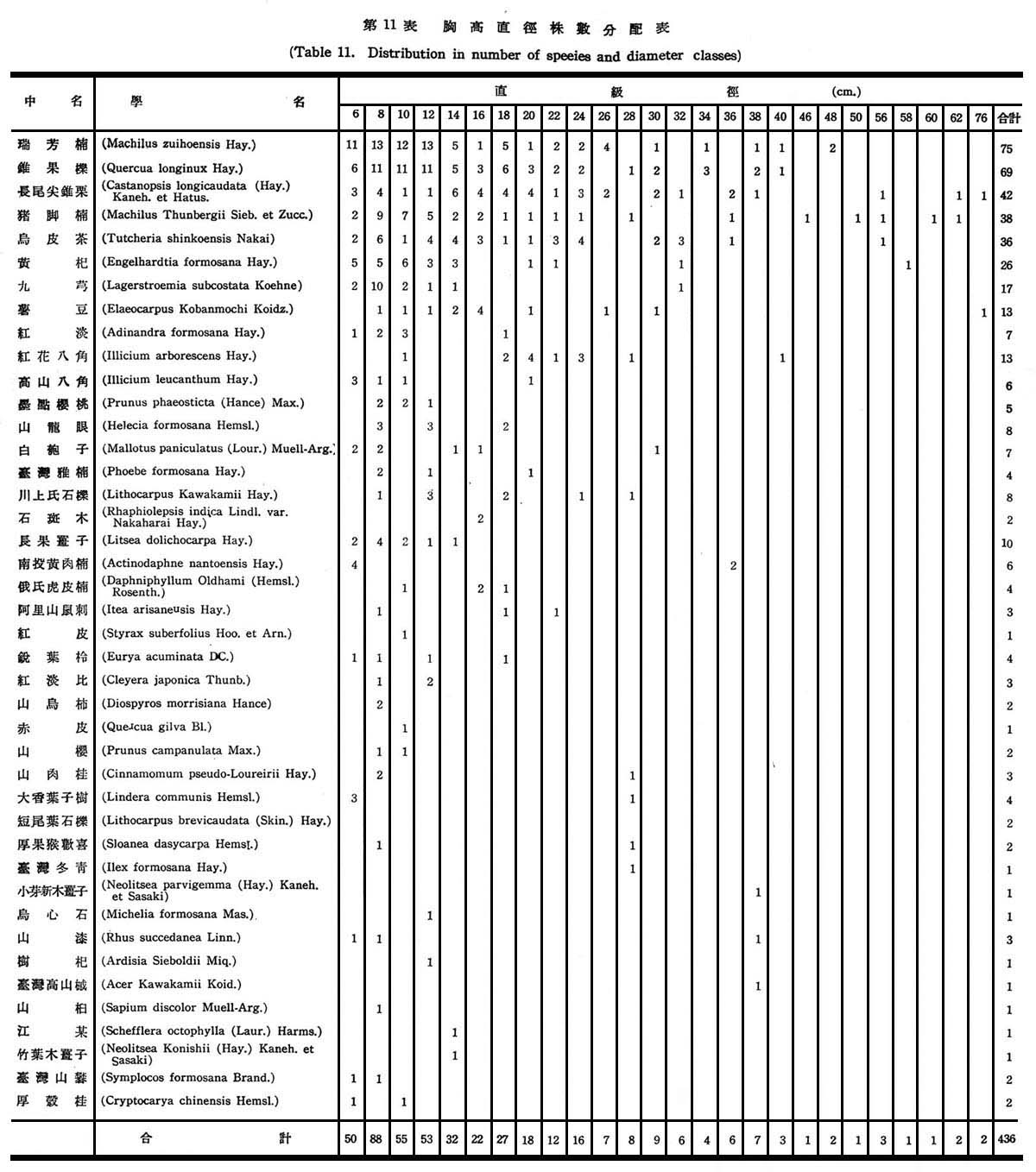
(3)胸高直徑與株數分配:
根據18個樣區測定結果,在目前狀態下亞熱帶降雨林之胸高直徑與株數分配如下圖所示:
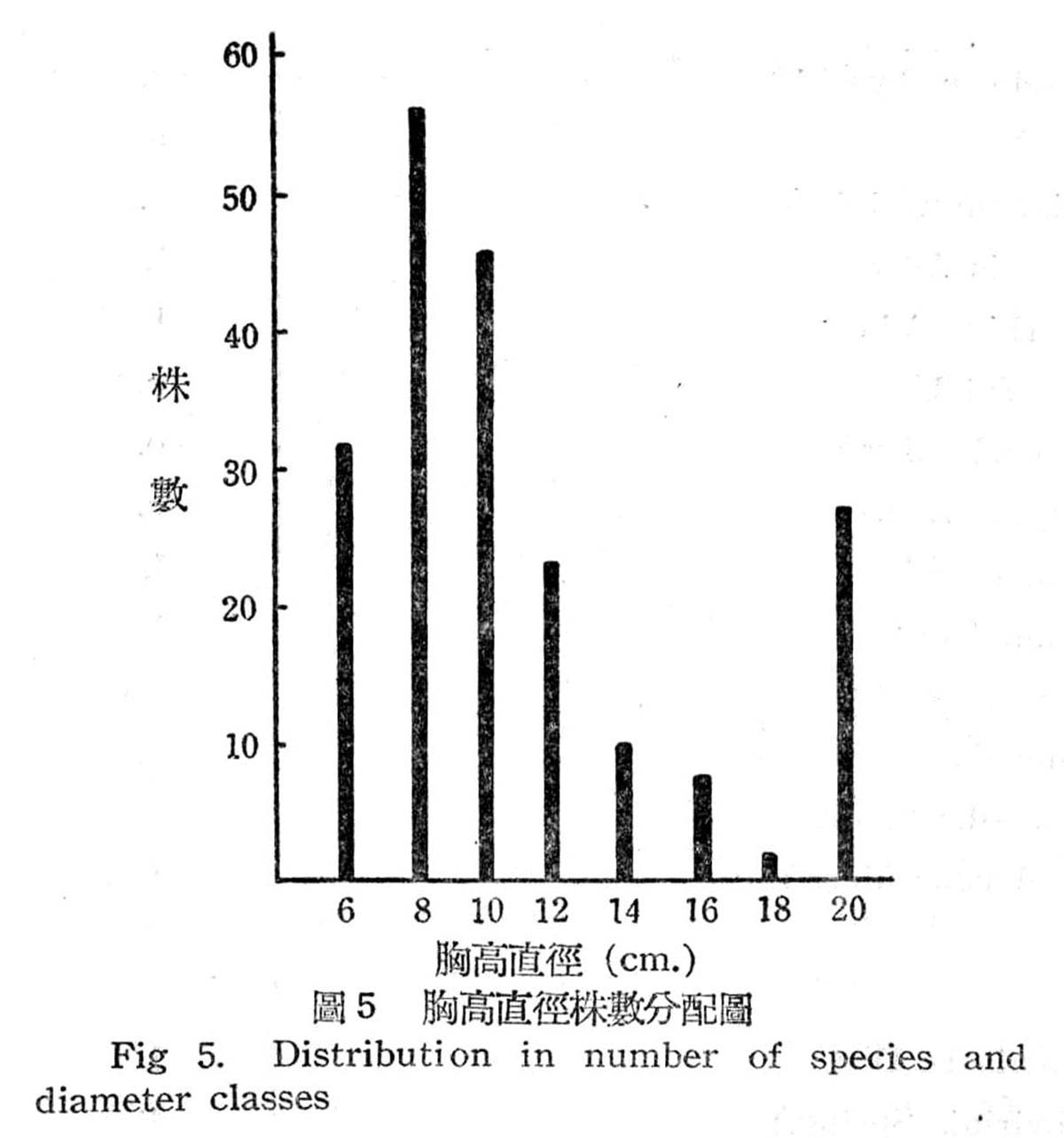
由圖5顯示此乃擇伐後之林型,蓋本省低海拔森林受人為破壞甚為普遍,直徑15cm以下佔77%,15~20cm佔23%,最大直徑未超過20cm以上者,故幼齡木佔絕大多數,壯齡木佔極少數,此森林早期已遭破壞,目前森林不過為伐採後之第二期幼年林。
(4)天然更新概況:(Natural Regeneration)
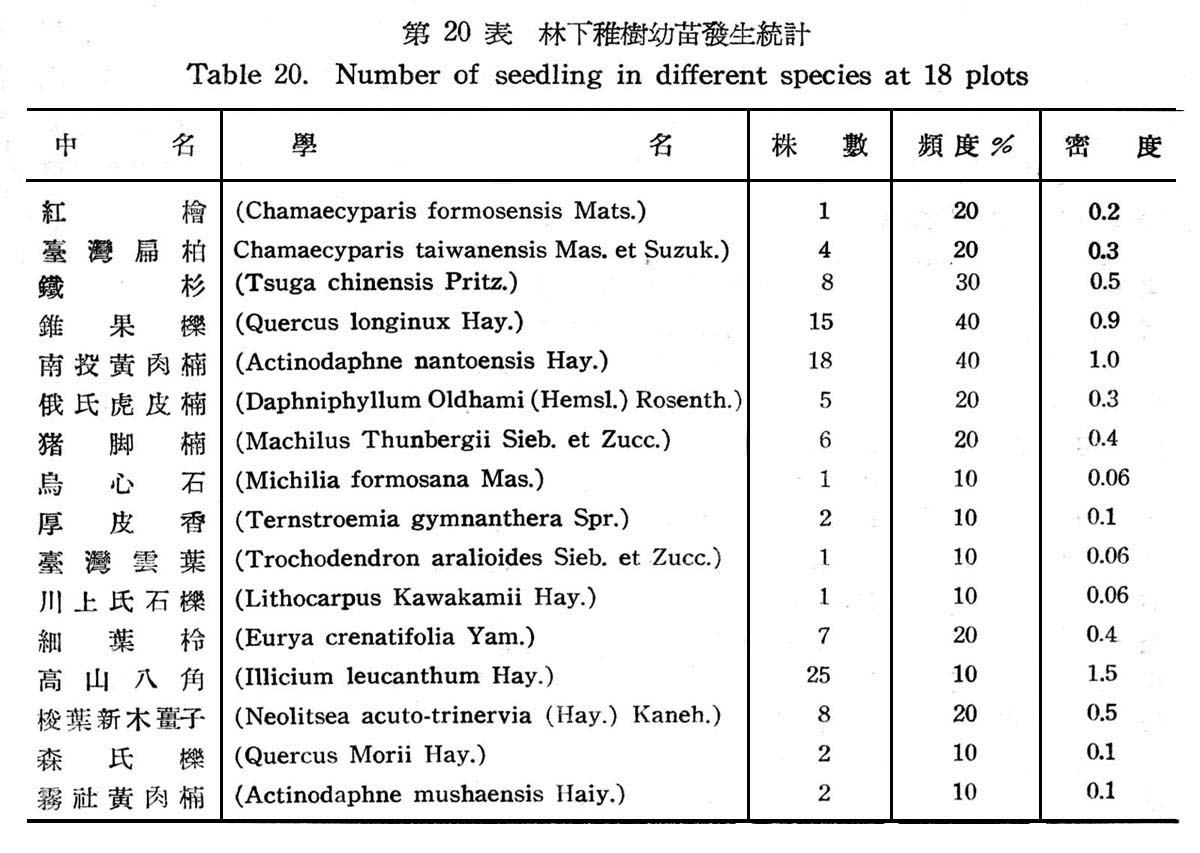
18f固樣區以測定其天然更新概況,以確定是否已達極盛點(climax.),結果如下表所示。
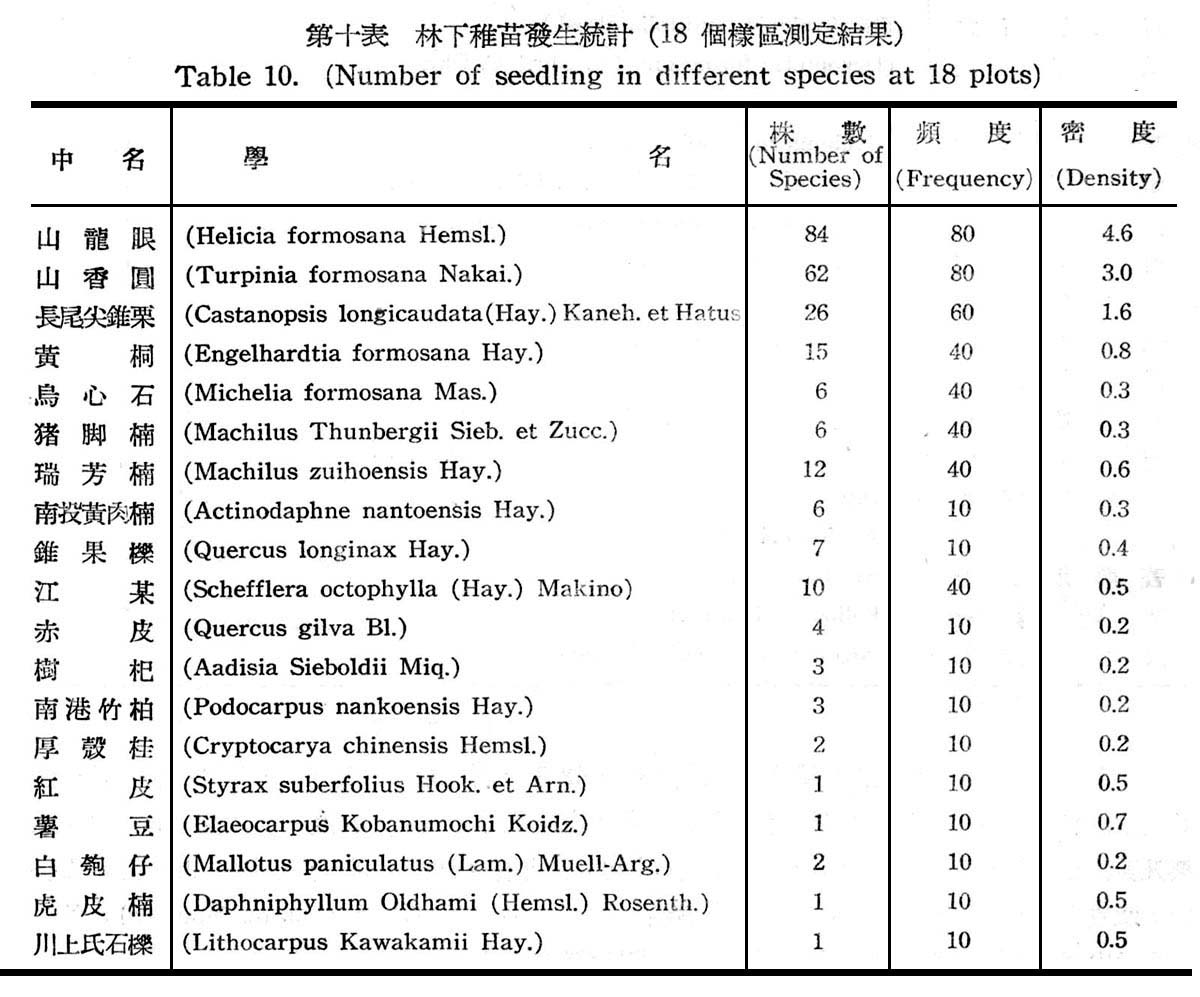
觀上表可知林下稚苗發生之頻度及密度,均以山龍眼及山香圓佔最大,其次則為長尾尖錐栗及黃杞,查山龍眼及山香圓均為亞熱帶降雨林習見之植物,近於半陽性者,但仍繼績延續而佔優勢,反之長尾錐栗及黃杞為以後侵入種近於中性植物,在第一層樹冠雖佔優勢,但其次代幼苗稚樹發生並不佔優勢,而較山龍眼及山香圓反居於劣勢,顯示長尾尖錐栗及黃杞,尚未達極盛植物社會(climax),此可確定仍在演變過程中,唯長尾尖錐栗及黃杞,將仍不失為該處之極盛植物社會主要份子。
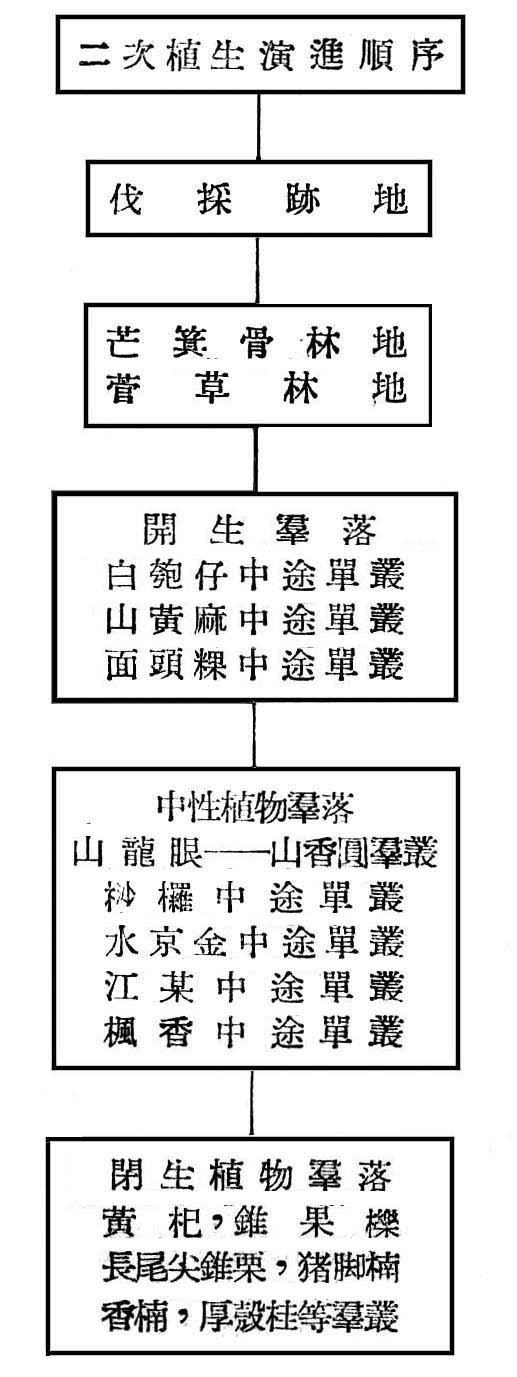
(5)生態演進之推論(Plant Succession)
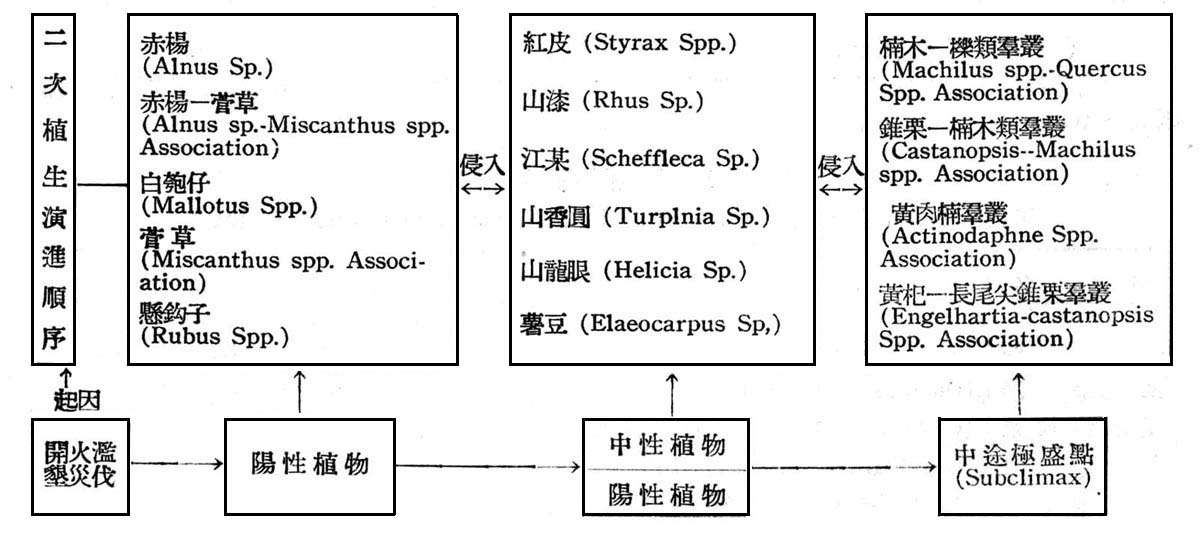
基於上列之分析,則大元山亞熱帶降雨林之生態演進,可作如是之推論,即大元山海拔高300~700m,亞熱帶降雨林早期受人為破壞,目前之第二期森林為伐採後發生之幼齡林,二次植生正在劇變(Secondary Succession),誘導二次植生演進之因子,不外乎濫伐,火災,開墾等因子所引起。原始植物演進Primary Succession)不外乎乾性植生列(Xerosere)與楊性植生列(Hydrosere),此地區植物生態之演進,應屬於乾性植生列殆無問題,且其演變過程中必須有一定之各時期(succession stage),且須較長久時間,至二次植物演進不須經一定時期與時間,當一已有森林之林地由於人為破壞與伐採後,林地暴露,土壤乾燥而固結,首先發生者為芒箕骨(Dieranopteris Spp.)及菅草(Miscantllus Spp.)植物群落所覆蓋,此等植物維持時間並不長久,即為開生群落(Open Community)如白匏仔(Mallotus Spp.),山黃麻(Trema Sp.),面頭粿(Glochidion Spp.)等極端陽性植物所侵入,此等陽性植物發育互相競爭結果,樹冠交叉而蔽蔭,反而造成不適於本身生存之環境,同時空氣中水分之下降,土壤由乾燥而濕潤,加以枯枝落葉之腐爛以增加土壤腐植質,立地略有改善,而介於中性與陽性植物間如山龍眼,山香圓,桫欏(Cyatllea-Alsophila Spp.),水京金,江某等相繼侵入,生長競爭結果,使陽陸植物逐漸淘汰,樹冠交叉更蔽蔭,土壤更為濕潤,日照減少,腐植質堆積,立地環境大為改善,則閉生群落(Close Community)如黃杞,錐果櫟,長尾尖錐栗,豬腳楠,香楠等次第侵入乃形成目前森林,在不受人為干涉情況下,目前森林將維持一較長久時期,唯根據現狀觀察知早期陽性植物雖逐漸減少,而介於陽性植物與中性植物間之山龍眼,山香圓,水京金等依然佔優勢,尤其次代之幼苗及稚樹頻度密度均佔最大,故此森林尚未達極盛點(Climax)。茲列一簡表以示其演變順序:
2.常綠闊葉樹林群系(Everngreen broad-leaved forest formation)
常綠闊葉樹林為本島主要植物群落型之一,樹種最為豐富,其分佈範圍恆在2500m以下,其中並有小部份為落葉喬木林外,主要以樟科(Lauraceae),殼斗科(Fagaceae),為主體,並參與若干山茶科(Theaceae),及山礬科(Symplocaceae)植物,在經濟價值上言似較亞熱帶降雨林高,大元山海拔高100~700m,列為亞熱帶降雨林已如上述,海拔高700~1300m則應列為常綠闊葉樹林帶,茲分述如下:
(1)群叢概述(Simple description of plant association)
瑞芳楠─錐果櫟群叢(Machilus zuihoensis-Quercus longinux, Association)
分佈於海拔高700~1300m,土壤濕潤肥沃山腹部佔最多,為構成大元山常綠闊葉樹林之主體,佔領面積廣潤,近於極盛植物社會。
長尾尖錐灤―豬腳楠群叢(Castanopsis longicaudata-Machilus Thunbergii, Association)
分佈於海拔高700~1300m,亦為構成大元山常綠闊葉之主體,土壤潤濕肥沃山腹佔最多,分佈面積亦廣,近於極盛極物社會。
南投黃肉楠一烏心石群叢(Actinodaphne nantoensis-Michelia formosana, Assocaiation)
分佈於海拔高700~1,300m溪谷向蔭濕環境中,土壤肥沃地多小團集狀態分佈,唯佔領範圍稍狹。
九芎群叢(Largerstromea subcostata, Association)
分佈於悔拔800~1,000m,近山腳溪傍甚為蔭濕之森林中,往往成小團集狀態,故其分佈範圍甚狹。
赤楊一管草中途群叢(Alnus formosana-Miscanthus sinensis, Associes)
分佈於海拔高700~1,000m,向陽而土壤略乾燥,火災後林地,崩壞地,溪傍或河床段丘地,多有發生,尤其崩壞地往往成大面積植物社會。
杜鵑―厚皮香群叢(Rhododendron Spp.-Ternstromia gymnantllera, Association)
分佈於山稜線部份,海拔高1,800~2,000m,陽光強烈,基岩暴露,土壤乾燥而貧瘠形成高山灌木林狀態,分佈範圍甚狹。
臺灣擦木─鹽膚木中途群叢(Sassafras Ramdaiense-Rhus javanica var. Roxburghii,Associes)
分佈於海拔1,200~1,500m,土壤乾燥陽光充足,開墾地或崩壞地,林綠部份往往成小團集狀態,分佈面積狹。
臺灣擦木一山胡椒中途群叢(Sassafras randaiense-Litsea cubeba, Associes)
分佈於海拔高1,000~1,300m,向陽乾燥土壤,往往成小團集狀態,以路傍開墾地,伐採跡地,露出地多有發現,分佈面積狹。
虎刺中途單叢(Damnacanthus formosanus, Consocies)
分佈於海拔高700~1,300m常綠闊葉樹林鬱閉咯破壞,而陽光折射林內略乾燥環境中,最習見之林下小灌木,多成小面積塊狀分佈,但範圍甚廣。
芭西木群叢(Barthea formosana, Association)
分佈於海拔高700~1,300m,亦為常綠闊葉樹林鬱閉良好之林下小灌木,但有時往往與虎刺混生成塊狀分佈,範圍亦廣。
玉山箭竹單叢(Pleisblastus niitakyamensis, Consociation)
分佈於海拔高1,000~1,300m,日照充足土壤乾燥環境中,伐採跡地,無立木地,往往大面積分佈。
菅草中途群叢(Miscanthus Spp., Associes)
分佈於海拔高400~1,800m,日照充足土壤乾燥貧瘠環境,崩壞地,開墾地,火燒地,溪傍河床地,均有大面積分佈。
塗氏苔團集(Carex Dunni, Clan)
分佈於海拔700~1,300m,常綠闊葉樹林鬱閉良好,土壤濕潤肥沃,最常見之林下地被植物,往往成小群落狀態,分佈範圍甚廣。
冷水花群叢(Pellionia scabra, Association)
冷水花─冷清草群叢(Pellionia Spp-Pilea Spp., Association)
分佈於海拔高700~1,300m,為常綠闊葉樹林林下主要地被植物,鬱閉良好,土壤深厚肥沃林地最為常見,有時成單叢,有時與冷水花(Pellinoia seabra),三萼冷水花(Pellinia trilobulata)阿里山冷清草(Piled brevicornuta)能高冷清草(Pilea nokozanensis)混合成群落分佈。
臺灣秋海棠單叢(Begonia laciniata var. formosana Consociation)
海芋單叢(Alocasia macrorrhiza, Consociation)
闊葉赤車使者群叢(Elatostema edule Association)
分佈於海拔高700~1,300m,常綠闊葉樹林林下習見之地被植物,蔽蔭土壤濕潤環境中成群生狀態。
紫苧麻團集(Villebrunea fruticosa, Colony)
分佈於溪傍,林緣,向陽而濕潤土壤成團集狀態,但分佈面積小。
(2)組成層次(Structure layers)
構成大元山常綠闊葉樹林組成層次30個樣區測定結果如下列各表所示。
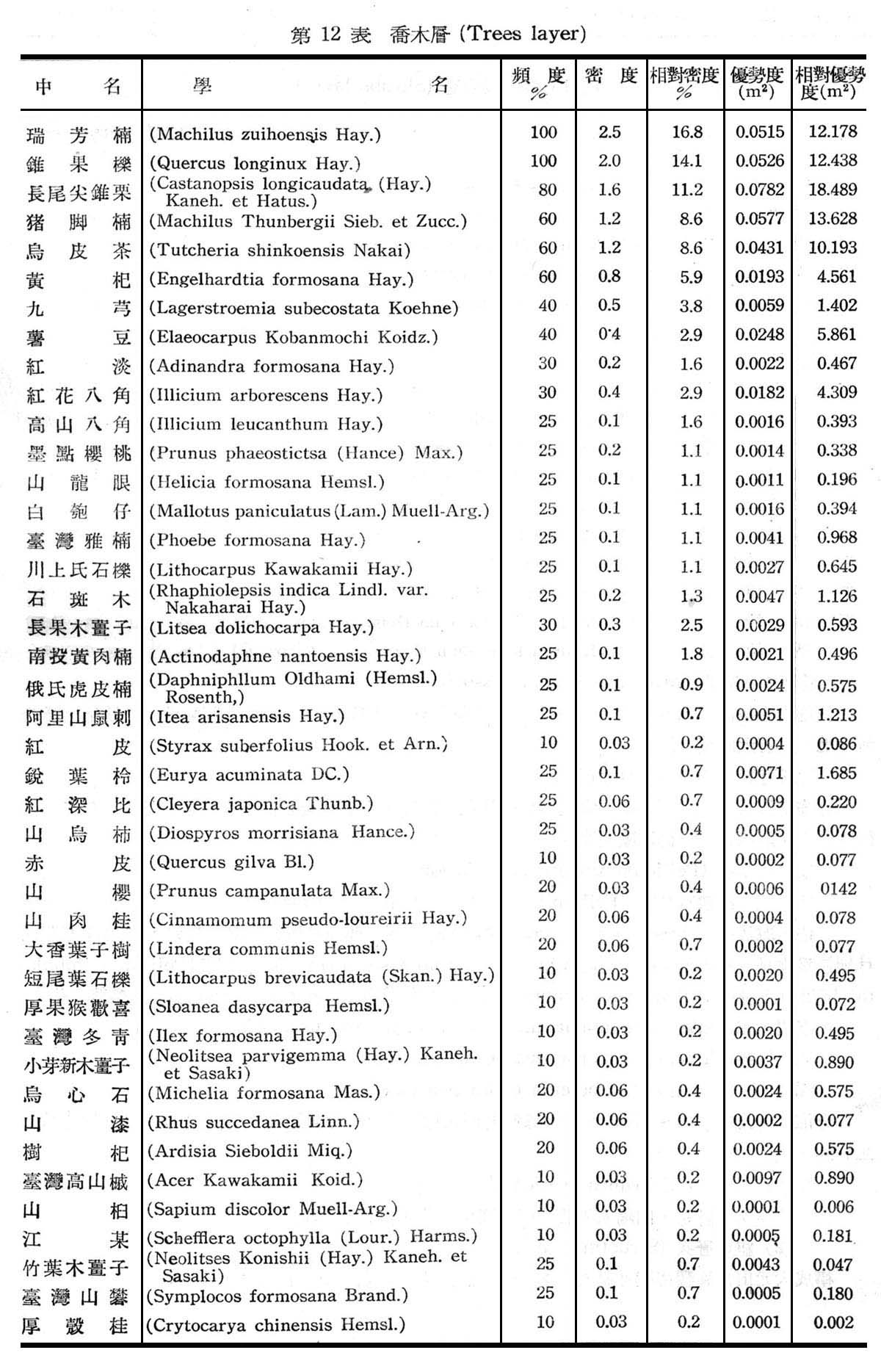


由上列12~14表統計可知構成大元山常綠闊葉樹料組成層次喬木層計42種,灌木層計36種,草本層計24種,羊齒植物計23種,苔蘚植物計5種,由第12表所示頻度,密度,優勢度等均以香楠,錐果櫟,長尾尖錐栗豬腳楠,烏皮茶佔優勢為優勢種(DominantsSpecies),黃杞,九芎,薯豆為次優勢種(Co-dominants Species),香楠,錐果櫟,長尾尖錐栗,豬肉楠為構成常綠闊葉樹林之主要群叢。
(3)胸高直徑與株數分配:
根據30個樣區測定結果,常綠闊葉樹林在目前狀態下,其胸高直徑與株數分配如第6圖所示。
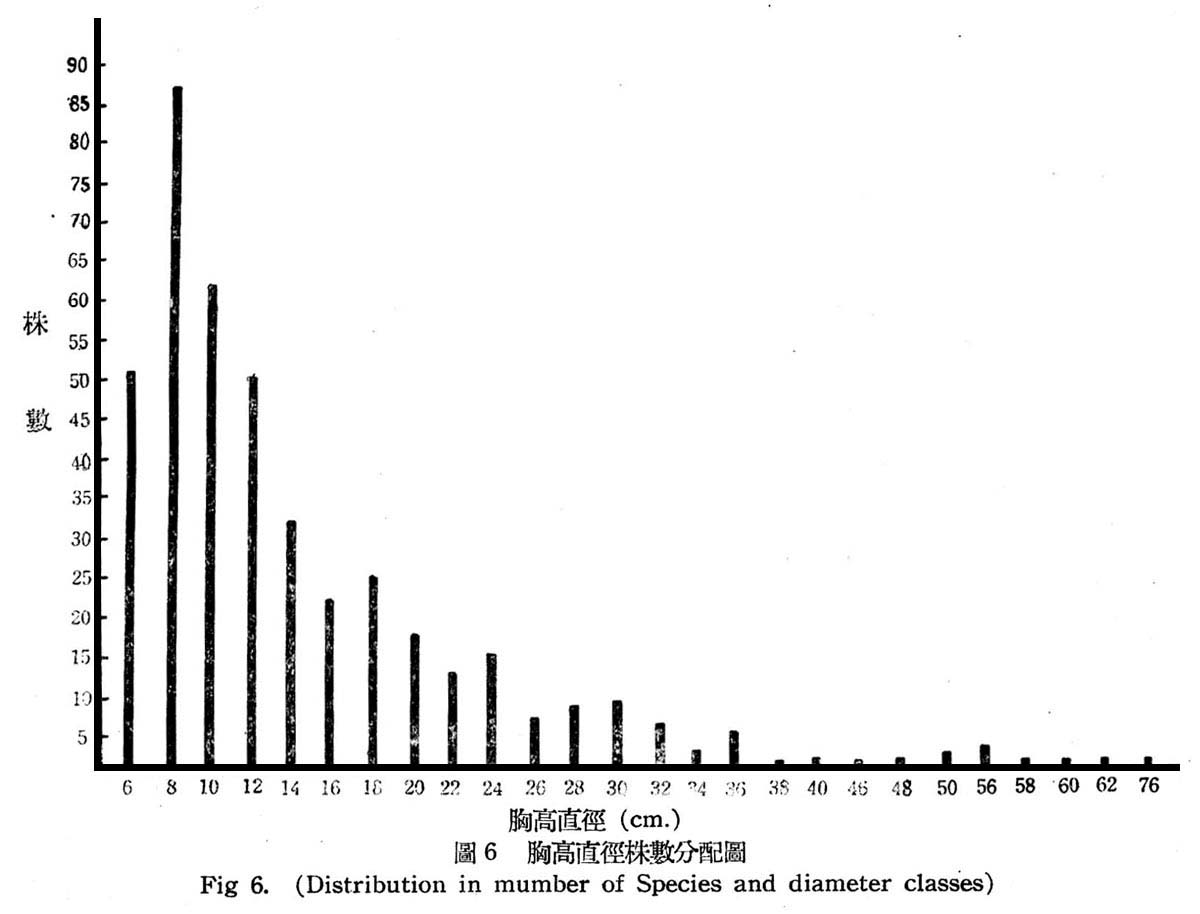
由圖6所示,大元山常綠闊葉樹林與下部亞熱帶降雨林之林型,大致相同,仍為早期受人為破壞與伐採,依目前現狀觀察仍第二期發生之幼齡林,胸高直徑12cm以下者占57.5%,12~38cm者占40%,40cm以上者占2.5%,即幼齡木占大多數,壯齡木占少數,老齡木幾已伐採殆億,此顯森林早期巳遭破壞。
(4)天然更新(Natural Regeneration)概況:
30個樣區以測定常綠闊葉樹林在天然情況下,其天然更新稚樹幼苗發生概況,以確定是否已達極盛點(Climax),結果如下表所示。
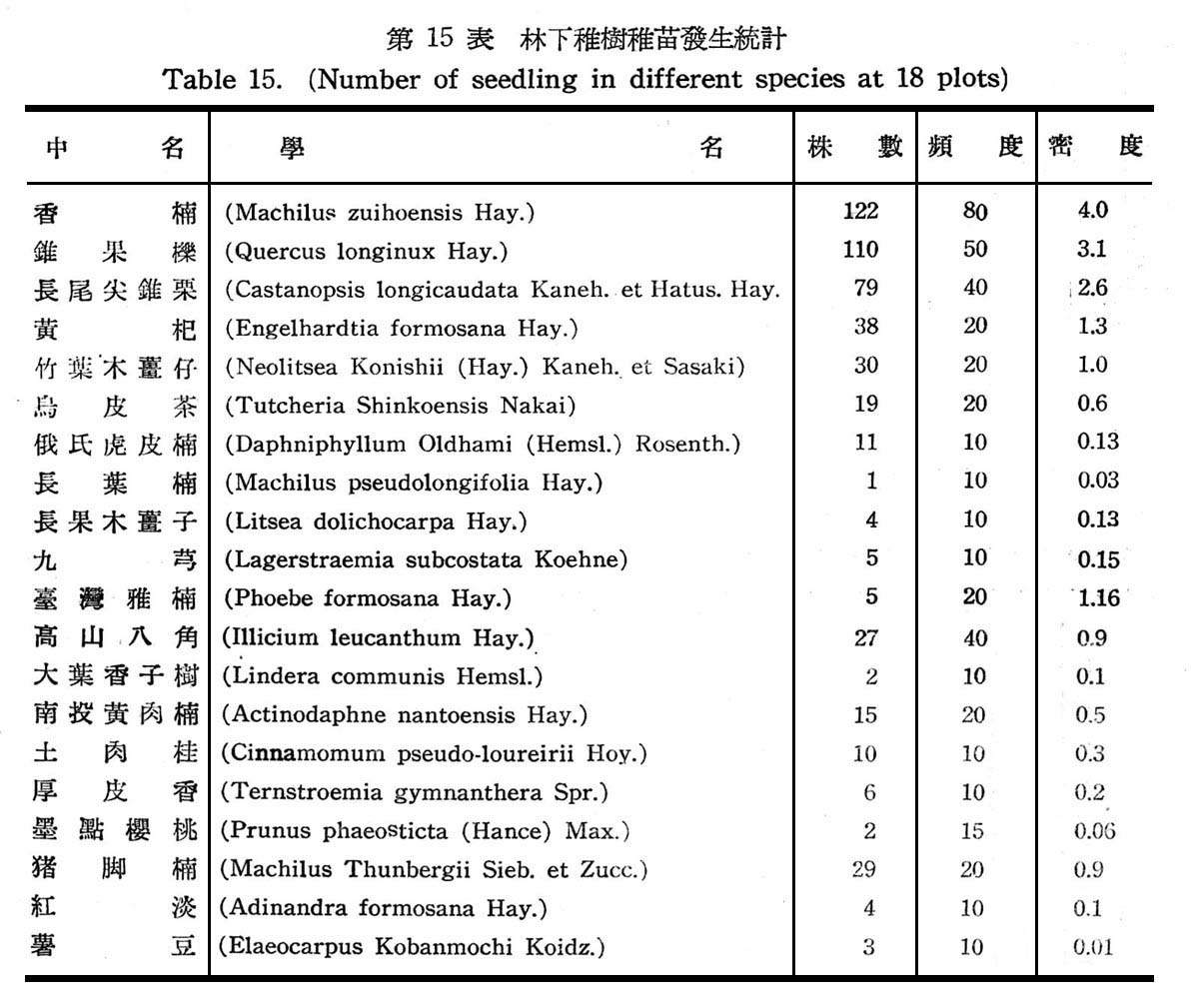
觀上表統計,香楠及錐果櫟不論在株數,頻度,密度上均占最大,其次則為長尾尖錐栗,豬腳楠,竹葉木薑仔,黃杞等:大致與喬木層之優勢種與次擾勢種相同,至其他稚樹幼昔均為本島闊葉樹林常見之植物,屬於陽性之稚樹幼苗已行絕跡,此顯示大元山常綠澗葉樹生態之演進已逐漸趨於極盛植物社會,此等值物將不失為該區之主要植物。
(5)生態演進之推論(Plant Succession)
依上述勸究結果,大元山常綠闊葉樹林生態之演進,吾人可作如是之推論,即大元山常綠闊葉樹林早期仍受人為破壞如伐採,火災,開墾等因子之影響,雖不若亞熱帶降雨林為害之烈,但林相巳遭受局部破壞,目前森林乃砍伐後發生之幼齡林,植被正在演變中,逐漸趨於極盛點(Climax),陽性植物社會早巳淘汰,現正由中性植物社會而進入較耐蔭植物社會中,即現有(1)香楠─錐果櫟群叢(2)長尾尖錐栗─豬腳楠群叢(3)南投黃肉楠─烏心石群叢(4)長尾尖錐栗一黃杞群叢等將逐漸發育旺盛,再查考其林下稚苗發生情況,仍為此四個群叢之主要植物,在不受人為干涉情況下,此四個群叢將維持大元山常綠闊葉樹林一較長久之時期,即現有森林非達安定點(Stabilization),不過為氣候控制下之極盛植物社會(Climax)發育過程中停留時期,反作用(Reaction)仍將繼續,在不受人為干涉狀態下發育大為旺盛,以改善立地環境,以適應更耐陰樹種之入侵。茲列 一簡表以示其演變順序:
3.常綠針闊葉混交林群系(Coniferous-evengreon broad leaved trees mixed forestformation)
大元山常綠針闊葉混交林自海拔1,300~2,000m,但實際上針葉樹在海拔高800m巳有發現,不過密度與頻度均小,故此林帶下部為闊葉樹林帶之延長,上部為針闊葉混交林帶,亦可稱為針葉樹與闊葉樹兩種不同植物社會之推移帶(Ecotone),針葉樹如紅檜臺灣扁柏不僅在密度大,且構成第一層樹冠為絕對優勢種,反之闊葉樹則為第二層樹冠成為被壓木,居於劣勢種地位。茲分述如下:
(1)針闊葉樹分佈概況:
植物分佈受溫度影響者大,一般闊葉樹所需溫度較高,且生育季節亦較長,因高溫對闊葉樹馴化較低溫為易,一年中有充足時問逐漸完成其生活週期(Life Cycle),供其發生新樹冠與葉簇,反之針葉樹所需溫度低,適宣於生育季節甚短之地區,因該地區無充足時問供其發生一新樹冠與葉簇,故多在高海拔與寒冷地區發現,故針闊葉混交林帶可代表該地區針葉樹與闊葉樹所需溫度兩者之極限,以大元山論紅檜雖在850m巳有發現,但究屬少數,而闊葉樹仍占優勢,海拔高1,300~2,000m臺灣扁柏,紅檜、鐵衫、香衫、亞杉等則為真正分佈地帶,有時幾成塊狀純林,而闊葉樹林反居劣勢。茲根據大元山分場不同海拔高之各林班每木調查資料統計結果如下表所示:
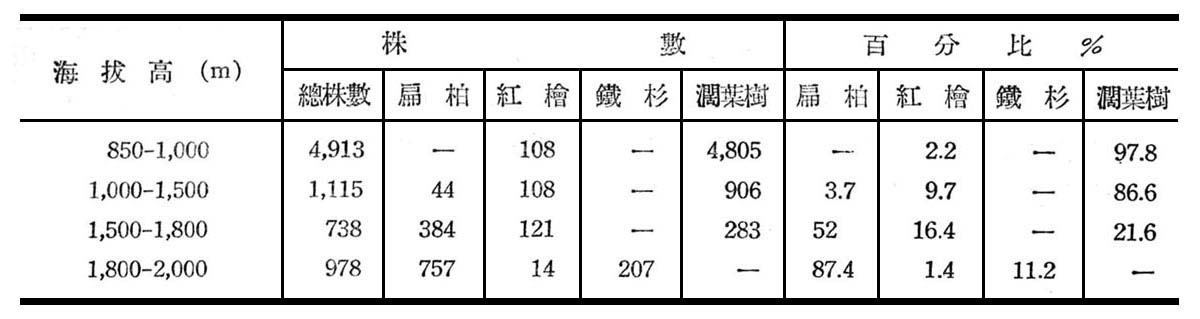
由上表顯示扁柏隨海拔高逐漸增高,紅檜則以1,500~1,800m處較多,潤葉樹則海拔愈低,分佈愈盛,鐵杉分佈僅限於1,800~2,000m左右。
(2)植物群叢概述(Simple description of Plant Association)
香杉群叢(Cunninghamia Konishii Association)
分佈於海拔高1,500m,早期可能有大面積之純林,或與紅檜扁柏成混交林,目前殆已伐儘,僅於伐採跡地保留有小面積之母樹林,或偶見與紅檜扁柏,鐵杉成混交之散生林,林下多玉山箭竹,菅草、懸鉤子(Rubus Spp.)等。
鐵杉群叢(Tsuga chinensis Association)
分佈於悔拔l,800~2,000m,接近於稜線尤多,有時成純林,但多與紅檜扁柏混交,林下植物為玉山箭竹,菅草,羊齒植物等,其第二層樹則為阿里山楠(Machilus arisanensis Hay.),高山八角(Illicium leucanthum Hay.),南投黃肉楠(Actinodaphne nantoensis Hay.),錐果櫟(Quercus longinux Hay.),森氏櫟(Quercus Morii Hay.)。
紅檜及扁柏群叢(Chamaecymris Spp. Association)
分佈於海拔高1,300~2,000m,為本林帶最主要之植物社會,以海拔高1,300~1,800m處最為旺盛,其分佈最低可下降至海拔高850m之闊葉樹林中常見有老木存在,有時成純林,有時為第一層樹冠之優勢種,其第二層樹冠則為樟柯─殼斗科群叢(Lauraceae-Fagaceae Association)即所謂照葉喬木林(Laurilignosa),林下植物多玉山箭竹,菅草、羊齒植物以緻脈鳳尾草足蕨(Plagiogyria euphlebia Association)群叢占最多,溪傍及崩壞地多見有小面積純林。根據大元山分場之統計及分佈現狀之觀察,扁柏在密度及數量上較紅檜多,故生產木材亦多。
樟科―殼斗科繁叢(Lauraceae-Fagaceae Association)
分佈於海拔高1,300~2,000m,以1,300m~l,7oom處最繁殖,為構成紅檜及扁柏林之第二層樹冠,主要植物為南投黃肉楠,錐果櫟,森氏櫟,長尾尖錐栗,山肉桂、阿里山楠、豬腳楠等所組成。
亞杉群叢(Taiwania Cryptomerioides Asssociation)
臆測早期有大面積分佈,此植物乃地質時代所遺留者,目前在大元為稀有植物,僅於翠峰梅拔高1,300處偶見一二株作為母樹者。
裡白瘤足蕨單叢(Plagiagyria euplilebia, Consociation)
分佈於海拔高1,300~2,000m,為針葉樹林主要地被植物群叢之一,此群叢大體上為陰性羊齒型,主林木伐採後多自行消減,繼起者則為懸鉤子(RubusSpp.)。
懸鉤子─菅草中途群叢(Rubus Spp.-Miscanthus sinensis, Associes)
分佈于海拔高1,300~2,000m,紅檜扁柏林伐採後,首先發生者為腺萼懸鉤子(Rubes glandufoso-calycinus Hay.),苦懸鉤子(Rubus indcesuaTh•)等常與菅草混合成塊狀分佈。
冷水花―冷清草群叢(Pellionia-Pilea Spp., Association)
分佈於梅拔高1,300~1,500m,為針葉樹林林下地被植物,於濕潤環境中每多發現,主要為冷水花(Pellionia Scabra Benth.),三萼泠水花(Pellionia trilobulata Hay.),阿里山冷清草(Pilea brevicornuta Hay.)等混合組,成成塊狀分佈。
芒箕骨―石松中途群叢(Dicroanopteris dichotoma-Lycopodium spp., Associes)
分佈于海拔1,500~1,700m,土壤乾燥向陽環境中,林緣,路傍每多發現。
赤楊中途單叢(Alnus formosana, Consocies)
分佈於海拔1300~2000m,崩壞地,傾斜地,溪傍,河床地每多發生,往往成大面積帶狀分佈。
擦樹一山胡椒中途群叢(Sassafras Randaiense-Litsea Cubeba, Associes)
擦樹―懸鉤子中途群叢(Sassafras-Rubus Spp., Associes)
分佈熊梅拔高1300~1500m,土壤略乾燥之開墾地,崩壞地,開墾地每多成小團集狀態。
菅草中途群叢(Miscanthus Spp. Associes)
分佈于海拔高1300~1500m,土壤略濕潤,溪畔、溪谷、河床地等環境中適宜於生長,成小面積分佈。
玉山箭竹單叢(Pleioblastus niitakayamensis, Consociation)
分佈於海拔1300~2000m,針葉樹林林下普偏大面積分佈,或針葉樹林伐採後即行侵入,雖生長稍差但其密度益增加,利用價值少,為造林撫育上之大害。
闊葉赤車使者一達令氏雙囊蕨群叢(Elatostema edule-Athyrium Doederleinii Association)
分佈於海拔高1,300~1,500m,溪傍濕潤土壤而蔽蔭環境中,為針闊葉混交林林下常見之地被物。此群叢屬于陰性者,良好鬱閉之常綠闊葉樹林之林下,更有大面積分佈。
苔蘚植物群叢(Moss-Pogantum Spp Association)
著生於岩壁空隙處,耐乾旱亦適於濕潤,此為乾性植生列(Xerosere)之開始,依抱子而傳播,根部可分越酸性以分解岩石使產生少量之土壤,以適應雜草之侵入。
(3)組成及層次(Structure layers)
大元山針闊葉混交林之組成分子及層次,由16個樣區測定結果如下列諸表所示:
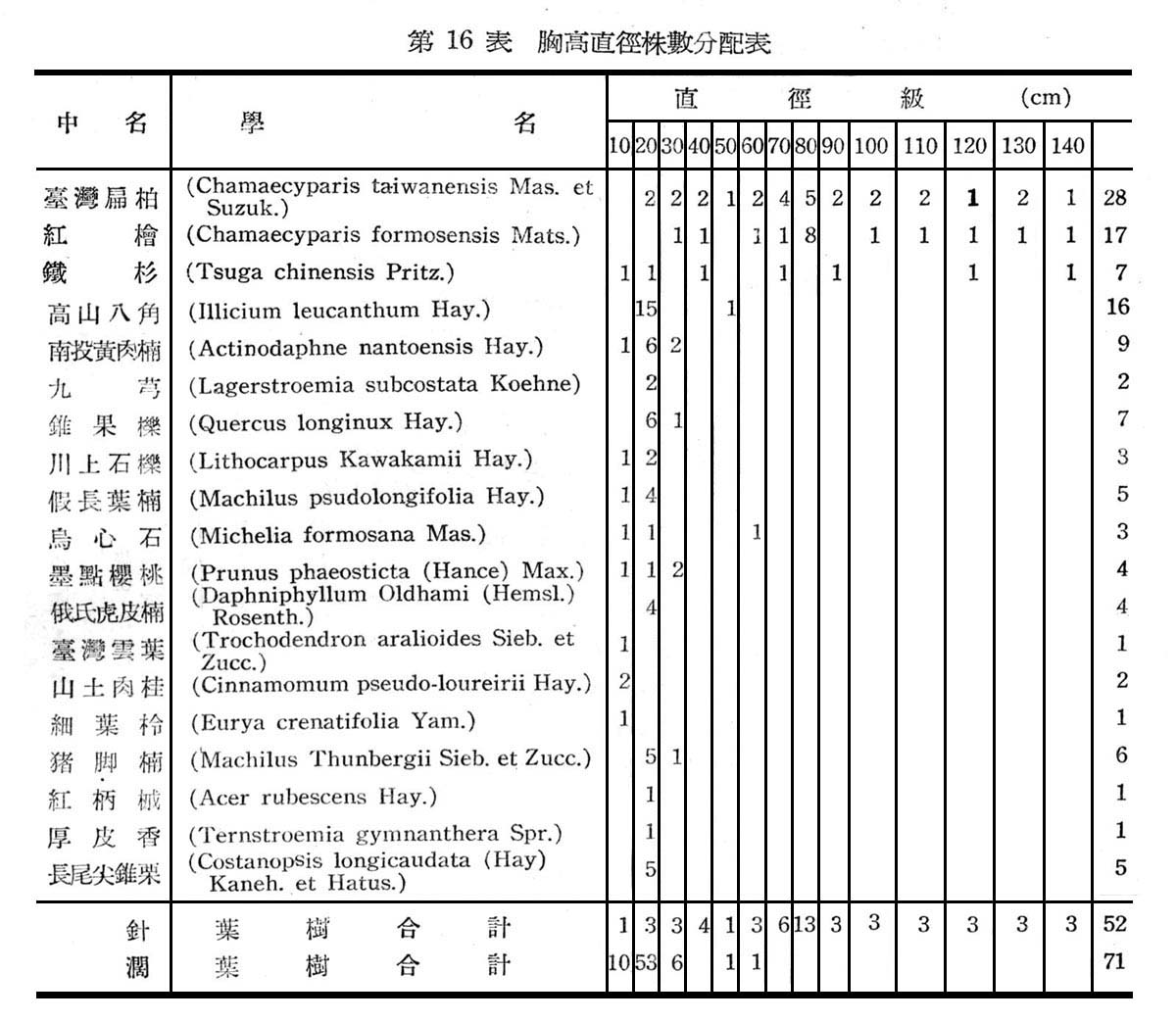
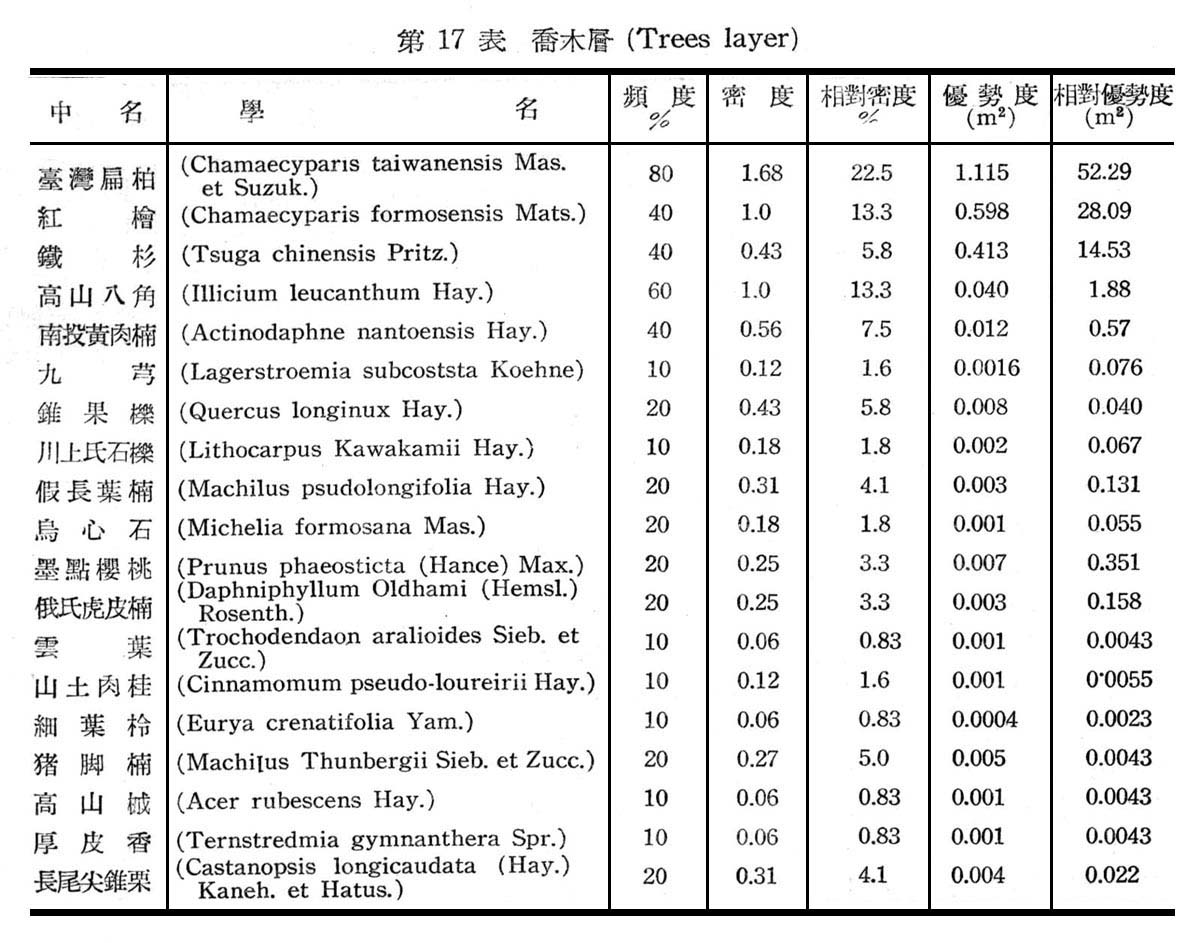
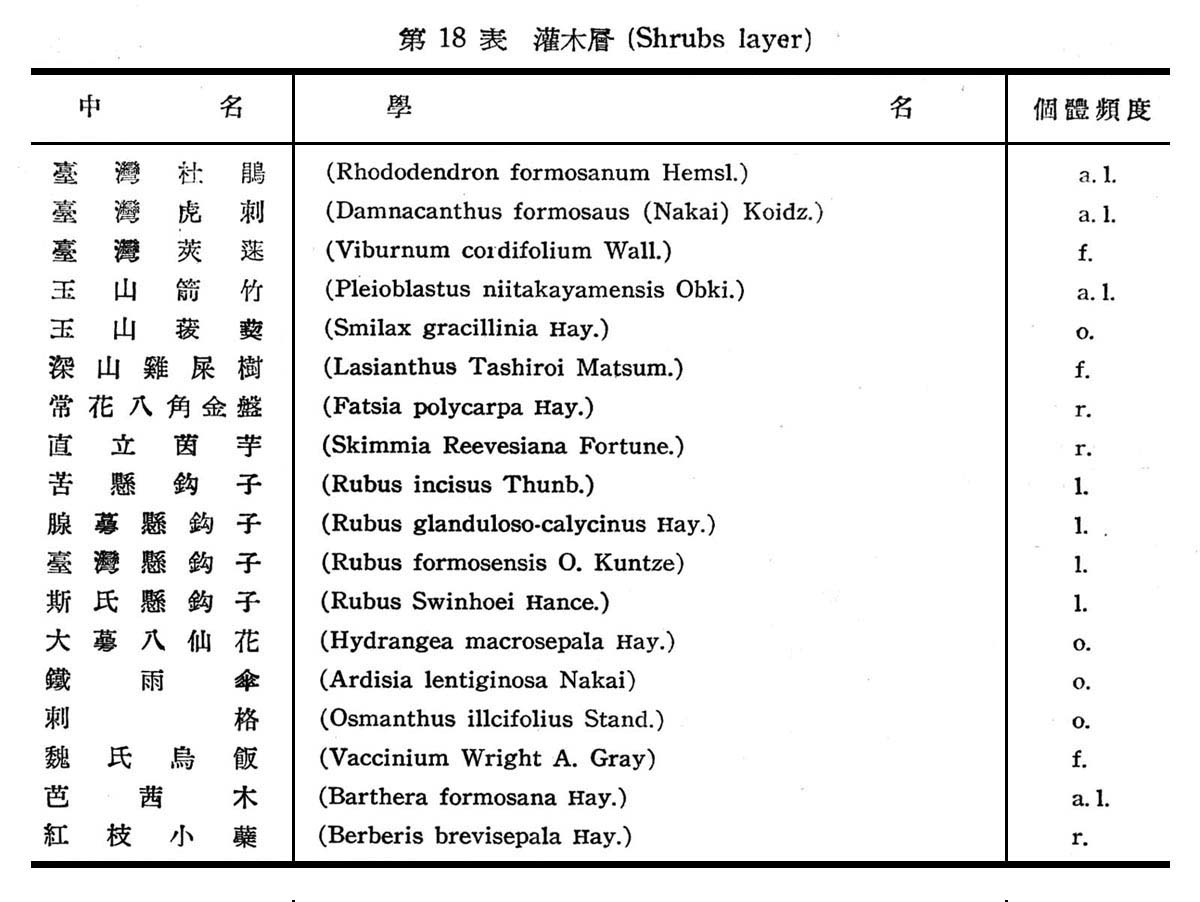

由表17一19構成大元山針闊葉混交林組成層次,計喬木層19種,灌木層18種,草木12種,羊齒植物17種,苔蘚植物4種,地衣1種。紅檜扁柏及鐵杉占絕對優勢,闊葉樹中高山八角在密度及頻度上雖大,但胸高斷面積甚小,故優勢度甚底,顯示闊葉乃後來侵入種成被壓狀態。
(4)胸高直徑與株數分配。
根據16樣區測定結果其在目前天然狀熊下,胸高直徑與株數分配如圖所示:
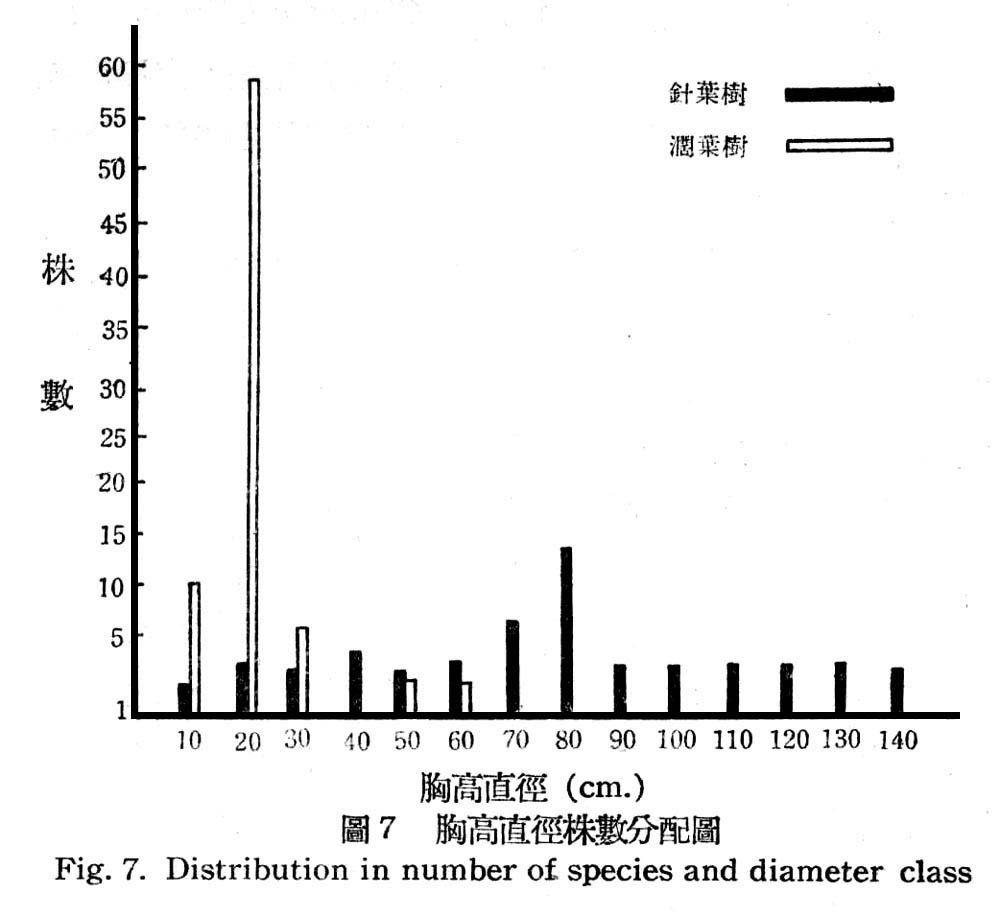
由圖7顯示闊葉樹幾全部為幼齡林,胸高直徑15~20cm占75%以上,極少數為壯齡林,反之針葉樹之林型為一齊林,即各直徑級分配頗為平均,即幼齡林,壯齡林,老齡林,三者均有平均分配,壯齡林稍多,故此早期尚未受破壞,就樹高言針葉樹平均高度均在20m以上,而闊葉樹平均高度均在15cm以下,顯示闊葉樹侵入後成壓被大。
(5)天然更新概況(Natural regeneration)
16個樣區測定其林下稚樹幼苗發生情況,給果如下表所示:
觀上表紅檜,臺灣扁柏,鐵杉在株數,密度,頻度均甚小,而稚樹幼苗發生多於崩壞地,路傍,溪畔,向陽地,枯倒木上,而在母樹林下反而少見,推究其原因,可能紅檜,臺灣扁柏,鐵杉等幼昔時期需要適當之日照,鬱閉良好之母樹下日照不足,又下層為玉山箭竹所密佈,種子難以直接著土,故稚樹幼昔均無法生長,是以在天然狀況下,針葉樹天然更新情況不佳,目前紅檜扁柏林雖然為氣候支配下之極盛植物社會(climatic Climax),恐將自行稍誠,至於闊葉樹林為針葉樹林鬱閉破壞後之侵入種,其中如高山八角,錐果櫟,南投黃肉楠等密度稍大,巳逐漸建立,目前雖為被壓木,生長不佳,如針葉樹砍伐後則將迅速占優勢,是以大元山紅檜扁柏林在目前狀況下,恐難以保持久遠,終將為闊葉樹林所取代。
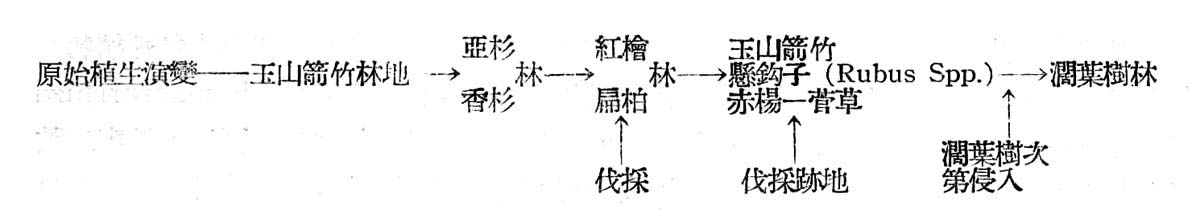
(6)生態演進之推論(Plant Succession)
探討大元山針葉樹林之起源,並非易事,臆測紅檜扁柏林發生必較闊葉樹早,然紅檜扁柏決非古時留傳迄今者,追溯古時之植物,不易搜集,故只有注意現代植物,容視其究自古代何種典型(type)留傳於今者,若針葉樹尚不失為古時典型種,此種植物之分佈情形可賽參老,臺灣之高山早期可能為玉山箭竹所占據,以後則有亞杉與香杉所侵入,此兩種植物均為古時地質時代遺留於今者,在大元山尚有少數分佈,香杉於紅檜扁柏林伐抖勁地保留有少數,或散生於紅檜扁柏林中,亞杉則更少見僅有一二株保留作為母樹林,故紅檜扁柏林或係由亞杉香杉林演進而來,由於亞杉及香杉不耐陽光照射,在天然狀態下更新復見困難,一旦林地部分遭受破壞,因而林相有空隙處,日照射入,遂變為較亞衫及香杉耐陽光之扁柏林,然扁柏奪紅檜耐陽性差,復轉變為較扁柏尤能耐陽之臺灣紅檜林之趨勢。至于闊葉樹乃因紅檜扁柏林鬱閉破壞後之林隙部份之侵入種,因而構成扁柏紅檜林極明顯之第二層樹冠,因受紅檜扁柏樹冠之抑制,故生長及品質均不佳,在不受人為干涉天然狀態下,現時成長之紅檜扁柏,乃氣候支配下之氣候極盛植物社會(Climatic Climax),然不能保持久遠,縱使現在森林,不施伐採終其天年,林相亦將有變遷,由於紅檜扁柏天然更新之不易,在現行砍伐細度下,首先侵入者將為王山箭竹,赤楊,懸鉤子(Rubus Spp.)菅草(Miscanthus Spp.)等,再後則為上述闊葉林次第侵入,競爭與反作用結果,將變為闊葉樹林,而臺灣扁柏與紅檜將不復存在,極少可保持現狀者。
茲列表以示其演變顯序如下:

六、指示植物之探討(Plant and Plant Communities as indicators)
植物或植物社會乃環境因子支配下之產物,某一環境下必發生某種植物或植物社會,其發生事例不但表示現在情形如乾濕肥瘠,作用(Proesses)如沖刺、伐木、火災、開墾等,用逮(Uses)如農地,牧地,林地等之指示物,以植物或植物社會對環境因子直接反應者,即所謂指示植物(Plant Indicator)。探討大元山之指示植物如山黃麻―榕群叢,山香圓―山龍眼群叢,木本羊齒之桫欏群叢,均為亞熱帶降雨林習見之氣候指標植物(ClimaticIndieator),尤其桫欏群叢可衡量亞熱帶植物垂直分佈之限界,常綠樹多於落葉樹乃表示該地區夏季與冬季均有適當之溫度與濕度,與豐富之降水。黃藤、拍拉木、葛藤、菊花木等藤本植物均為熱帶性氣候下之產物,乃表示長熱之夏季,適度之冬季適當之溫度,與豐富之降水。在一個大植物社會中,局部現出小團集之特殊植物群落者,如芒箕骨群叢發現乃表示該土地為乾燥貧瘠近於酸性土壤者。海芋(Alocasia macrorrhiza Schott.),秋海棠(Begonia Spp.),泠水花(Elatostema Spp.)等地被植物發生區域,乃表示該局部地區環曉蔽蔭土壤濕潤者。櫟類(Quercus Spp.)發生多表示土壤濕潤深厚而肥沃者,白抱仔,山胡椒,鹽膚木,擦木局部發生地區乃指示從前已有森林由於火災,開墾,砍伐而森林捎失者。臺灣赤楊發生地區乃山崩後之餘物。極端陽性植物連續發生者乃表示該地區植生初期正在演變中即所謂連續指標植物(Serial Indicator),亞杉香杉乃地質時代留傳於今者,此可指示森林變遷上之參考。蘚苔植物大面積發生於林內乃指示森林濕度高與降雨多,玉山箭竹發生地乃表示過去為針葉樹林地因伐採而消失者。凡此種種均可指該地區之環境一般與所發生之事列。
七、植物生活形(Life from of plants.)
植物生活形為一區域內氣候狀態之指示奢,對植物社會與地理上之被院甚為重要,生活形分類式各學者分類式各有不同,其中以Raunkaier氏對於高等植物生活形分類式最為適用,其分類式如下:
(S)Stem-succulents多肉莖植物
(E)Epiphytes著生植物
(MM)Mesophanerophytes大形喬木
(M)Microphanerophytes喬木
(N)NanoPhanerophyes灌木
(Ch)Cliamaepllytes地表植物
(H)HemicryptoPhytes半地中植物
(G)Geoph外es地中植物
(HH)Helophytes&HydrophyteS水生植物
(Th)Thyerophytes夏生一年植物
茲將大元山採得植物目錄列表如下,並冠以簡單符號以表示其生活形。

由上表計79科,170屬,263種,其中地衣1種,苔蘚5種,羊齒植物50種,顯花植物207種,其生活形統計如下列所示:
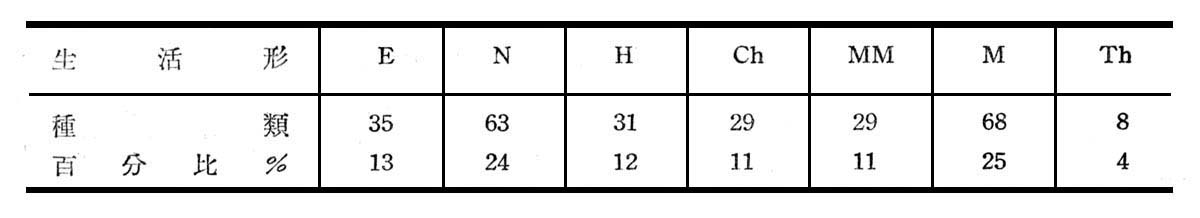
由上知(N)灌木24%,(M)喬木25%,MM大形喬木11%合計地上植物占60%,此顯示大元山為地上植物氣候(Phanerophytes Climate)。
八、結論(Conclusion)
一、就經濟價值言,海拔高500m以下,亞熱帶降雨林,林業經濟價值最為低微,海拔高500~1,300m,常綠闊葉樹林帶次之,海拔高1,300~2,000m為最高,本省亞熱帶降雨林帶,由於人口稠密,濫伐,開墾等現象甚為普遍,森林破壞最烈,大元山亦不例外,屬於有用林木既少,胸高直徑一般均在20cm以下,海拔高500~1,300m以上之常綠闊葉樹林帶,以樟科及殼斗科植物,為常綠闊葉樹組成之主要分子,雖有相當數量之分佈,唯多屬於幼齡林,其胸高直徑大部份均在20cm左右,目前似乎未達開發利用時期,林業經濟價值目前亦不高,海拔高1,300~2,000m,主要為紅檜,扁柏林,生長良好巳達伐採時期,蓄積豐富,為本省北部主要木材生產地,其中以扁柏較紅檜多,本林帶林業價值最高。
二、綜合研究結果,各林帶之植物社會其環境因子略有差異,就氣象上言,大元山屬於重溫濕氣候,一年中有適當之溫度及濕度,與豐富之降水,宜於林木之生長,海拔高800m上下,即亞熱帶降雨林之上部,與常綠闊葉樹之下部,年平均溫度為攝氏16.5度,海拔高1,300m常綠闊葉樹林帶,年平均溫度為攝氏14.8度,最高溫度為八月份攝氏27.1度,最低溫度以二月份攝氏4.2度,年平均溫度為87%,年總雨量為5,6092m.m.,月年平均雨量為467.4m.m.,海拔高1,900m針闊葉混交林帶林年平均溫度為攝氏10.9度,最高溫度為八月份攝氏24.1度,最低溫度以二月份攝氏零下4.8度,年平均濕度為94%,年總雨量為3,869.7m.m.月平均雨量為323.00m.m.一般言之,闊葉樹所需之濕度低於針葉樹,而溫度則恰好相反,因而闊葉樹所需溫度較高,生育時間亦較長,針葉樹生育時間較短,所需溫度亦較低,故針葉樹每生長於寒冷地區,海拔高800~1,300m闊葉樹林帶每升高100m溫度遞減率為攝氏0.37度,海拔高1,300~1,900m針闊葉混交林帶每升高海拔l00m溫度遞減率為攝氏0.60度。就土壤上言,大元山土壤為粘板岩及頁岩風化而成,有效深度均在lm上下,土壤深淺肥瘠不一,一般言之,傾斜度大者土層淺,傾斜度緩和者土壤深,土壤多呈濕潤狀態,含水量均在50~60%,通透性良好,闊葉樹土壤多屬于酸性,針葉樹土壤多呈酸性或強酸性。
三、大元山之林型可分為(一)亞熱帶降雨林:海拔高700m以下,受人為破壞最烈,屬於第二期森林,依目前森林現狀及次代稚樹幼昔發生統計上觀之,多屬於半陽性植物,有用植物不多,此森林如欲改善其林相,除非以人為力量加以促進之,在天然狀態下林相之改變,恐須長久時間,反不如加以皆伐以重建人工林為佳,(二)海拔高700'1300m,常綠闊葉樹林帶,林相尚佳,觀其組成分子,尚不乏有用樹種,且次代之幼苗稚樹發生統計上觀之,亦多為較耐陰樹種,目 前雖為幼齡林,如善用人力促進,以改善其林相,即採用適當之擇伐,以促進上木與下木迅速生長,不須甚長時間,林相當可改觀,此森林仍屬有希望。(三)海拔高1300~2000m,主耍為針葉樹之紅檜,扁柏林,闊葉樹為針葉樹鬱閉破壞後之侵入種,成為極明顯之第二層樹冠之被壓木,在直徑與高度上相差懸殊,唯紅檜與扁柏在天然默態下,天然更新情況不佳,推究其原因可能為(1)良好鬱閉下日照不足,(2)林下什草繫生,(3)玉山箭竹密佈下種子無法著土,此等因予均足以影響種子發芽與稚苗生長,目前紅檜扁柏林乃氣候支配下之極盛植物社會(Climax),由於天然更新之不易,即不加伐採,終其天年,恐亦難保持其久遠,終將為闊葉樹林所取代,唯紅檜扁柏並非必定無法維持,例如在崩壞地,溪畔,路傍,火燒地,枯木上及什草稀少地上,仍見有稚樹幼苗之出現,且生長亦佳,此顯示如能善用適當方法以人力促進之,以造成適于幼苗發生之環境,則紅繪扁柏林,仍可延績。
四、大元山之氣候與土壤大致已如上述,針葉樹以1,300~2,000m處生長為最佳,山中腹部多紅檜,愈上部則扁柏較多,700m~1,300m為闊葉樹林分佈最旺盛地帶,700m以下為亞熱帶降雨林,是以林帶之區分已甚顯明,則造林實施亦應循此天然林帶,選鐸適當樹種實施造林,但應注意者:
(1)臺灣赤楊為崩壞地常見之樹種,效果甚佳,故大元山崩壞地舊造林可採用之。
(2)針葉樹林砍伐後應立即造林,否則玉山箭竹甚易侵入,將來實施迭林將倍感困難。
(3)福衫,柳杉造林均須濕潤肥沃土壤,應注意局部土壤之選擇,因大元山林地土壤均不甚深厚也。
(4)香衫與亞杉應注意採種與育苗,並實施造林以保存此優良樹種。
(5)實施針葉樹單純造林,不如實施混交林,不違背天然法則。
(6)根據調查觀察結果,維持紅檜扁柏今後之延續,採用天然更新方汰,應已不成為問題,但尚須進一步之實驗與研究。
九、附圖(Photograph)
章樂民所附為黑白圖片,加以民國50年印刷技術設備尚差,影像模糊不易辨識,為讓讀者容易閱讀,盡量選用適地適時的彩色圖片補述。上圖是原來附圖,下圖是補述的彩色圖檔。


圖8 南澳北溪兩岸之闊葉樹林之林相景觀
Fig.8.Physiogonomy of evengreen broad-leaved forest along river coast on Mt.Ta-yuan. At l,000m. above the Sea level.


圖9 針闊葉混交林之林相景觀上部近稜線多針葉樹中下部為闊葉樹林
Fig.9. Physiogonomy of Mixed forest formation on Mt.Ta-yuan. At 1500m. above the sea level.


圖10 伐採跡地所見之紅繪扁柏林
Fig.10. Chamaecyparis for Mosanensis and Chamaecyparis Taivanensis in cut over land on Mt.Ta-yuan. At 1300m. above The sea level.


圖11 針葉樹林林相之景觀
Fig.11. Physiogonomy of Coniferous foreston Mt.Ta-yuan. At l,400m. above the level.


圖12 溪流兩傍發生之赤楊群叢
Fig.12. Alnus formosana Association commonly growing Along the bank of riverat 750m Above the sea level.


圖13闊葉樹林下部之桫欏群叢
Fig.13. Alsophila pustulosa-Cyathea Taiwaniana commonly forming an Associ ation in subtropical rain forest on Mt.Ta-yuan at 700m. above the sea level.


圖14 北向之闊葉樹林內細齒樹苔滿掛于樹幹顯示林內濕度甚大
Fig.14. Pilotrichopsis dentata forxning an Association, in Evengreen broad-leaved forest at 1500m. above the sea level.


圖15針闊葉交林帶之闊葉樹林之林相較高大者為針葉樹之紅繪,扁柏,鐵杉,細小位於下方者為闊葉樹林
Fig.15. A Mixed forest formation on Mt.Tayuan. first layer: chamaecyparis formosensis, chamaecyparis taiwanensis and Tsuga chinensis.
Second layer: evengreen broad-leaved trees.


圖16常綠闊葉林帶鬱閉略破壞發生之台灣擦樹群叢
Fig.16. Sassafras randiense forming an Association, on Mt.Ta-yuanat 1600m. above the sea level.


圖17 針闊葉混交林帶之鐵杉群叢
Fig.17. Tsuga chinensis forming an Association, on Mt.Ta-yuan at 1800m. above the level.


圖18 針闊葉況交林帶之紅檜扁柏群叢
Fig.]8. A Purestands of chamaecyparis Spp. Association on Mt.Ta-yuan. At l,60Om above the sea level.

圖19 針闊葉混交林帶之臺灣杉
Fig.19. Taiwania cryptomeriedies, a very rare species on Mt. Ta-yuan. At l,300m. above the sea level.


圖20 針闊葉混交林帶之香杉群叢
Fig.20. Cunninghamia Konishii Association,
Growing in cut over land on Mt.Ta-yuan at 1650m. above the sea level.


圖21 亞熱帶降雨林破壞後之菅草林地上方為赤楊(Alnus formosana)侵人,中間為八字桫欏(Alsophilla pustulosa)侵人,下方為菅草(Miscanthus spp.)
Fig,21. A grass land (Miscanthus spp.), are invaded by Alnus formosana and alsophilia pustulosa. On Mt.Ta-yuan at 700m. above the sea level.


圖22 常闊葉樹林帶向陽地常見之火母炭藤群叢
Fig.22. Polygonum chinense Association, an under growth in evengreen broad leaved foreston Mt.Ta-yuan at 1200m. above the sea level.


圖23大元山常見之菅草群叢
Fig.23. Miscanthus spp. Association, in upland meadow on Mt.Ta-yuan at 1000-1500m. above the sea level.

圖24 亞熱帶降雨林之上部,常綠闊葉極林之下部,常見之地被植物,闊葉赤車使者群叢
Fig.24. Elatostema edule Association, anundergrowth in evengreen broad-leaved forest on Mt.Ta-yuan at 800~15O0m. above the level.


圖25亞熱帶降雨林之上部,常綠闊葉樹林之下部,常見之地被植物,海芋群叢
Fig.25. Alocasia macrorrhiza Association an under-groth in subtropical rain forest at 500~1000m. above the sea level.


圖26常綠闊葉樹帶常見之地被植物─台灣秋海棠群叢
Fig.26. Begonia laciniatvar. Formosana Association, an undergrowth in evengreen broad leaved forest at 800m. above the sea level.


圖27 亞熱帶降雨林之上部,常綠闊葉樹林之下部之紫苧麻群叢
Fig,27. Villebrunea fruticosa Association, an under growth in subtropical rain forest at 700m. above the sea level.


圖28 亞熱帶降雨林之上部,常綠闊葉樹林之下部之達令氏雙囊蕨─闊葉赤車者群叢
Fig.28. Elatostema edule-Athyrium Doederleinii Association, an under growth in subtropical rain forest at 800m. above the sea level.


圖29 紅檜扁柏林下之玉山箭竹群叢
Fig.29. Pleioblastus nittakayamensis Association, an undergrowth in Comiferous forest occurs a large area on Mt.Tayuan at 1300~2000m.
Above the sea level.


圖30 紅檜扁柏林伐採跡地之玉山箭竹群叢侵入
Fig.30. Pleioblastus nittakayamensis Association, an invasion species, in cut over land of chamaecyparis formos Anensis and chamaecyparis taiwanensis.


圖31 針葉樹伐採後初期發生之陽性灌木─腺萼懸鉤子等
Fig.31. Rubus glandulasocalycinus-Rubus spp. Association in cut over land of coniferous forest.


圖32 闊葉樹林內之著生植物─孔雀蕨
Fig.32. Epiphytes(Asplenium Seropenduform)


圖33 闊葉樹木內之著生植物─飛刀劍與書帶蕨
Fig.33. Epiphytes (Cyclo-Phorus Lingua and Vittaria suberecta)


圖34闊葉樹林枝之著生植物─山蘇花
Fig.34. Epiphytes(Asplenium antiquun.)


圖35針葉樹林內之著生植物─膜蓋蕨
Fig.35. Epiphytes(Leucostegia parvipinnata)


圖36 針葉樹林內之著生植物─孔雀蕨屬
Fig.36. Epiphytes (Asplenium Sp.)


圖37 著生於樹幹上葉狀地衣
Fig.37. Epiphytes (Foliose lichensp.)